
場景一:燈光幽暗的臥室,一名俊秀的男子正躺在床上。一名漂亮的年輕女子穿著睡衣走近床前。鑽石婚戒在她的左手上閃閃發光,她的右手緊抓著一條小小的藍色紙帶,俯身親吻男人的耳朵。
她:“親愛的,現在時候正好!”
場景二:同一臥室,同一對人,顯然正在做愛,但細節都被幽暗的燈光恰到好處地遮蓋了。鏡頭搖到一本日曆(顯示時間的流逝),一隻戴著同一鑽石婚戒的纖手慢慢地翻著。
場景三:同一對漂亮夫婦,充滿幸福地抱著一個清爽的微笑著的嬰兒。
她:“親愛的,我真高興,排卵試紙能告訴我們什麼時候正好!”
最後一個畫面:同一纖手的特寫,抓著一條小小的藍色紙帶。丈字說明:“排卵試紙,家居測尿,排卵即知。”
假如狒狒能理解我們的電視廣告,它們一定會認為這個廣告太小題大作。無論雌狒狒還是雄狒狒都無需一種激素測試用品來檢測雌性的排卵期,即雌性卵巢排出一個卵子時可以受精的期間。相反,此時雌狒狒的陰部隆起,變成鮮紅色,在一段距離內均可看見。雌狒狒還散發出一種獨特的氣味,如果愚笨的雄性狒狒還不開竅,雌狒狒就會蹲伏在它面前展示臀部。多數其他雌性動物同樣能察覺自己的排卵期,並會同樣勇敢地用視覺信號、氣味或者行為通知雄性。
我們認為臀部鮮紅的雌狒狒很奇特。實際上,我們人類是那種在動物世界中為數很少的排卵期難於察覺的生物之一。男人沒有可靠的方法確知他們的性伴侶何時能受精,傳統社會中的女性同樣無能為力。我知道許多女性在一次月經週期的中間部分會經歷頭痛或其他感覺,可是,如果不是科學家們告訴她們,她們不會知道這些就是排卵期的徵象——即便是科學家也是直到1930年前後才搞清楚這一切的。同樣,儘管可以教會女性通過觀測自己的體溫或粘液來找出排卵期,但這和雌性動物擁有的本能仍有極大的不同。如果人類也能有這樣的本能,那生產排卵期測試用品和避孕用具的廠家就不會這樣生意火爆了。
人類幾乎持續不斷的性行為也是非常怪異的,這種行為可以被視作人類隱蔽的排卵期的直接後果。多數其他動物都將性活動限定在廣而告之的排卵期前後的發情期階段。英語名詞estrus(發情期)和形容詞estrous(發情期的)都源於希臘詞語的“牛虻”一詞。牛虻是一種追逐牛群並將牛群驅趕得發瘋的昆蟲。在發情期,一頭雌狒狒在l個月的禁慾後要交配多至100次,一頭雌性巴巴裡獼猴甚至平均17分鐘交配一次,對種群中的每一頭成年雄性至少施惠一次。一夫一妻的長臂猿伴侶無性生活長達幾年,直到雌性給它最幼的孩子斷奶後再次發情。一旦雌性再次懷孕,長臂猿伴侶恢復禁慾狀態。
可是我們人類在成年後的任何一天都可以有性生活。女性可能在任何一天需要性,男人性行動時不會挑剔他們的性伴侶是否處在排卵期或能否受孕。幾十年的科學調查認為,女性對性的興趣確有週期性的變化,但是女性在週期的哪一階段對男性的性舉措最有興趣仍未能確知。因此,人類多數的交合發生在女性並不能受孕的時刻。我們不僅在這個週期中錯誤的時刻性交,甚至在孕期內和顯然不可能懷孕的絕經期後發生性行為。我的許多新幾內亞朋友覺得應當經常做愛直到孕期結束,因為他們相信精液反覆注入有助胎兒身體發育。
按照天主教教義,性的功能等同於生產。從“生物學”的觀點來看,人類的性確實是精力的極大浪費。為什麼女性不能像其他多數雌性動物一樣顯示出明確的排卵徵兆,這樣我們就可以將性限制在對人有所裨益的時刻。本章力求能揭示隱蔽性排卵的進化、女性對性幾近持續的接受能力和性娛樂——這三位一體的作為人類性活動核心的怪異的繁衍行為。
現在,你一定認為我是那種無事生非,找些問題來闡述一下的脫離現實的科學家的典型。我聽到世界上幾十億人都在抗議:“沒有問題要你解釋,除了為什麼賈裡德·戴蒙德(Jared Diamond)是這樣一個大白癡。你難道竟會不理解為什麼我們總是需要性嗎?當然是因為樂趣!”
不幸的是,科學家們不滿足於這個答案。當動物從事性行為時,從它們的激情投入看,似乎它們也在享受樂趣。如果說長達12小時的交配時間是某種證明的話,那麼袋鼠看來享受著比人類多得多的樂趣。那麼,為什麼多數動物只在雌性可以受孕時才會想到性的樂趣呢?正如解剖結構在自然選擇中發展變化,行為也同樣在發展變化著。因此,如果性是享樂,必然起因於自然選擇。性對狗來說確實也是樂趣,但只有在恰當的時刻才是:狗同其他多數動物一樣,養成了在性能有所成果的時刻享受性的良好習慣。自然選擇惠顧那些能以自己的行為將基因傳遞給最多後代的個體。但是,如果你瘋狂到在不可能有孩子的時候享受性,自然選擇怎樣幫助你養出更多的孩子來呢?
一個最簡單的例子,可以描述多數動物物種性活動的目標取向性,這就是我在第二章探討過的鳥類花斑鶲。通常一隻雌性花斑鶲只在產卵前幾天做好受精準備時才要求交配;當開始產卵時,她對性的興趣便蕩然無存,抗拒雄性的性要求,對雄性的態度與先前大相逕庭。鳥類學家們做了一個試驗,在產卵結束時將其配偶轉移,讓20只雌性花斑鶲寡居,但試驗者們觀察到其中有6只花斑鶲在2天內試圖與新的配偶交配,3只實際進行了交配,也許還有更多不為人知地完成了這一切。顯然,雌鳥企圖迷惑雄鳥以使其相信它們是能受孕的。當卵終於孵化時,雄性不可能知道這些雛鳥實際上另有生父。至少有幾例,這種花招成功了,雄鳥如同生父一樣擔當起哺育雛鳥的職責。因此,這個試驗絕不能被認為顯示雌鳥作為快樂的寡婦,追求純粹的性的愉悅。
人類在排卵期隱蔽、隨時可接受性和性娛樂這些方面的獨樹一幟,只能是因為人類是以這種方式進化的。但對於智人——唯一有自我意識的物種來講,特別反常的是女性居然無法意識到自己的排卵期,而其他雌性動物,即便蠢笨如母牛,都對此一清二楚。這裡肯定有某些特殊需要來掩蓋聰明的女性的排卵期。如同我們將要看到的,科學家們在試圖弄清這些特殊情況的因果關係時遇到了出乎意料的困難。
為什麼其他多數動物很明智地吝惜交配的資源?理由很簡單:性太花精力、時間,並且有傷亡危險。我們來列舉一些理由說明為什麼你不應該無謂地與愛人做愛:
1.雄性精液的產生代價高昂,以致發生基因突變後精液產量降低的蠕蟲,其壽命要比正常的蠕蟲來得長一些。
2.性佔用了時間,而這些時間本可以用於覓食。
3.兩性交合時,有受驚和為肉食動物或敵人捕殺的風險。
4.老年人有可能因為性的緊張而疲累身亡:法皇拿破侖三世在性交時中風,納爾遜·洛克菲勒在性交時猝死。
5.雄性動物間爭奪發情的雌性不僅會傷害雄性,而且時常嚴重傷害雌性。
6.許多動物,包括人類還有在婚外性行為時被抓的風險。
因此,如能擁有其他動物一樣的性效率將是人類的一大進步。人類從自身明顯的低效率中得到了什麼補償性的收益呢?
科學思考更趨向集中在人類另一個不同尋常的特點:人類的幼兒在很多年裡依賴於父母的呵護,而多數哺乳動物的幼崽斷奶後即開始自己覓食,很快變得完全自立。因此,多數雌性哺乳動物能夠並且確實獨自養育幼崽,無需雄性協助,雄性對於雌性只有在交配時才是必不可少的。然而,對於人類來講,多數食物需要複雜的技術手段才能獲得,這是蹣跚學步的嬰兒無論是在技能上還是在智力上都無法企及的,由此人類的孩子至少在斷奶後10年內仍需由他人提供食物,父母雙方共同養育孩子無疑比一方獨力承擔要容易得多。即便在今天,單身母親養育幼兒仍是辛苦備至,更不用說在人類只是採集者和狩獵者的史前時代。
現在我們來設想一下,一個處於排卵期的、剛剛受孕的穴居女性的兩難窘境。對於其他哺乳動物,雄性在交配後很快就會離開,尋找另一個處於排卵期的雌性交配。而對於穴居女性來講,男性的離開意味著她生下的孩子將瀕臨飢餓或者乾脆就是被謀殺。那麼,為了拴住男性她能做些什麼呢?聰明的方案是:保持對性的接受力,即便是在排卵期後!隨時滿足他交配的需要。這樣,男人就會被吸引住,無需尋找新的性伴侶,甚至甘願讓她分享狩獵所得。因此,性娛樂被假定具有維繫人類配偶共同養育後代的紐帶作用。實際上這是人類學家早就接受的理論,並且有許多可圈可點之處。
然而,由於我們對動物行為瞭解得越來越多,我們意識到這種性愛鞏固家庭的理論仍然有很多問題無法解釋。黑猩猩尤其是倭黑猩猩雖然比我們人類更頻繁地性交(多達每天數次),可它們仍然亂交,並不存在配偶契約。相反,我們可以列舉無數哺乳動物中的雄性陪伴伴侶和後代並不是出於性的誘惑。長臂猿實際上是一夫一妻的,經年累月過著無性的生活。你向窗外看去,可以觀察到雄燕雀是何等勤勉地協助伴侶餵養雛鳥,而性在雌鳥受精後已經停止了。即使是一夫多妻的雄性大猩猩一年當中也只有幾次性交機會,他們的配偶總是在哺乳或者不在發情期。那麼,為什麼女人必須經常以性籠絡男人,而其他雌性動物則無需如此呢?
人類配偶和那些禁慾的其他動物物種間有一個重要的區別,那就是長臂猿、多數鳴禽和大猩猩是散居的,每對(或者每群)配偶各自佔有領地,這種居住模式幾乎沒有邂逅潛在的婚外性伴侶的可能。而傳統人類社會的典型特徵也許就是配偶與其他配偶們結群生活,彼此必須為經濟目的而合作。要找出一種近似的平行居住生活的動物來,我們必須超越人類的哺乳動物親戚,而將視線投到高密度築巢群居的海鳥身上。但海鳥的夫妻們,也不像人類那樣為經濟目的彼此依賴。
於是人類性的兩難窘境就是,父母必須長年共同哺育不能自立的孩子,同時抵擋周圍其他富有生育能力的成年異性的誘惑。婚外性行為是破壞婚姻並危及父母哺育孩子的合作關係的幽靈,它始終在人類社會遊蕩。不管怎樣,人類進化發展了隱蔽的排卵期、恆久的性的接納力、共同哺育後代以及通姦的誘惑等特點。那麼它們是怎樣磨合的呢?
科學家們過去對這些矛盾的評價產生了許多相互對抗的理論,每一種都反映著它的倡導者的性別。例如,有一名男科學家推出賣淫理論:女性操淫業是為了換取男性狩獵者的捕獲物。還有一個男科學家提出私通優化基因理論,推斷如果一個穴居女性不幸被其氏族嫁與一個無能的丈夫,她可以憑借其持久的對性的接受能力吸引一個有優秀基因的鄰近穴居男人並由此婚外受孕。
同時,一個女科學家提出了反避孕理論,她非常清楚,新生兒與產婦的體形之比,比我們的近親猿類大得多,以致人類女性的生育極其痛苦和危險。一名體重約45千克的女性通常生下2.7千克重的嬰兒,而體形2倍於她的雌性大猩猩(90千克)生養的卻是一半大小的幼崽(1.35千克)。結果,人類女性在現代醫學誕生之前常常死於生育,並且女性在分娩時需要人幫助(在現代發達國家是產科醫生和助產上,在傳統社會則是接生婆和年長婦女);而雌性大猩猩分娩時則無需幫助,從無困生育而死亡的記錄。因此根據反避孕理論,某些穴居女性清楚生育的痛苦和危險,也清楚她們的排卵時間,並不適當地運用這一知識,以達到避孕目的。這樣的女性無法將她們的基因代代相傳,以致這個世界上充斥著對自己的排卵時間一無所知的女性,並因而無法在易受孕時避免性交。
在這一片氾濫的解釋隱蔽排卵期的假設中有兩個假設,即“居家父親”理論和“多父”理論所起來似乎最有道理。有趣的是,這兩個假設是完全相反的。“居家父親”理論假定隱蔽排卵期的進化是為發展一夫一妻制,迫使男人留在家庭,強化男人對其妻子所生育子女盡父親的責任。“多父”理論則假設隱蔽的排卵期可以使女性擁有眾多的性伴侶,而使眾多的男人無法確知自己是否繁衍了她的孩子們。
首先來看一下“居家父親”理論,它由密歇根大學的生物學家理查德·亞歷山大(Richard Alexander)和凱瑟琳·努南(Katharine Noonan)提出。為了理解他們這一理論,我們先來想像一下,如果女性如同雌狒狒一樣用鮮紅色的臀部來公開她們的排卵期,那麼婚姻生活是何等情狀。丈夫會從她妻子臀部的顏色準確無誤地識別出她排卵的日子,然後在這一天會留在家裡,奮力做愛以使她受孕,傳遞基因。而在其他時候,從妻子蒼白的臀部他會明白與她做愛毫無用處。於是他會出去遛躂,尋找別的無人衛護的紅臀女士,以使她們受孕,遺傳更多自身的基因。他心安理得地將妻子留在家中,因為他知道她此時不會接受男人的性要求,並且無論如何也不會受孕。這就是雄鵝、雄性海鷗和花斑鶲的所作所為。
對人類來說,這種公開排卵期的婚姻生活無疑有著可怕的後果。父親將很少在家,母親不能獨力養育孩子,嬰兒將大批死去。對於父母雙方這都是糟糕透頂的事,因為誰也不能順利地傳遞基因。
現在我們來描繪一下相反的情形。丈夫對妻子的受孕時間無跡可察,於是他為了更多一些機會令其受孕,不得不留在家中,在每個月中盡可能多地與她做愛。迫使他留在家中的另一動機是為了時刻保衛妻子,防止別的男人乘虛而入,因為她很可能恰好在他出門的某一天懷了孕。如果某天晚上妻子恰好排卵,而花心的丈夫不幸正躺在別的女人的床上,那其他男人很可能就會在這位花心丈夫的床上令他妻子受孕,而丈夫卻在別處與另一無望受孕的女人通姦,浪費精液。在這種截然相反的設想中,由於男人無法識別哪些鄰人的妻子可以受孕,他就缺少理由出去遛躂。結果皆大歡喜:父親們在家忙碌,共同照料孩子,嬰兒茁壯成長。父母雙方都由此得益,因為他們可以成功地傳遞基因。
實際上,亞歷山大和努南強調的是由於人類女性獨特的生理狀況迫使丈夫們留在家中(至少要比他們本來停留的時間長)。女性因為招募到一個積極的育兒夥伴而受益;而男性如果肯與妻子合作,遵循妻子身體變化規律辦事的話,他亦因之受益。男人一直留在家中,便有信心認為他正在共同養育的孩子確確實實攜有他的基因。他無需恐懼在他離家狩獵時,他的妻子(如同一隻雌狒狒的作為)會亮出鮮紅的臀部將自己的排卵期廣而告之,吸引成群的求偶者,和周圍的每一個男人公開交合。男人對這些基本規律如此深信不疑,以致即使他們明知不能令妻子受孕,他們仍會在孕期和絕經期後與妻子繼續交合。因此,在亞歷山大和努南看來,女性發展隱蔽的排卵期和持久的性的接受能力是為了促進一夫一妻制、共同育兒和加強父親們盡父道的信心。
和以上觀點相對抗的是由加州大學戴維斯分校的人類學家薩拉·赫爾迪(Sarah Hrdy)提出的“多父”理論。人類學家很早就認識到殺嬰現象在許多傳統人類社會是普遍存在的。儘管現代各國已制定法律制止這一現象,可直到赫爾迪和其他學者著手這一領域的研究前,動物學家對於在動物中殺嬰現象的發生率並無估算。現在記錄在案的有這一現象的動物物種除從獅子到非洲獵犬等一大批物種外,還包括人類的最近親:黑猩猩和大猩猩。成年雄性對於未與其交配過的雌性所生的幼崽尤其容易做出殺戮的行徑。例如,雄性入侵者力圖取代原來的雄性並奪取其雌性配偶群,奪位者無疑知道所殺的幼崽並非親生。
自然,殺嬰現象令人驚恐,我們不禁要問為什麼動物(及早先的人類)會頻繁地做出這種勾當來。經過仔細思考,結論是謀殺者可以據此得到令人髮指的遺傳優勢。雌性在哺育幼崽的時候不可能排卵。但是殘忍的入侵者和它剛剛接管的種群中的幼崽沒有絲毫的血緣聯繫,殺掉這樣一頭幼崽後,它可以終止幼崽母親的哺乳期,刺激它恢復發情週期。在許多或者說是多數動物殺嬰和奪位情況下,殺嬰者使承受喪子之痛的母親受孕,使其生育攜有殺嬰者基因的幼崽。作為幼崽死亡的一個主要原因,殺嬰現象是動物中母性面臨的嚴重進化問題,它們因此喪失其在被殺後代身上的遺傳投資。例如,由於奪位的雄性大猩猩企圖接管配偶群而殺嬰,使得雌猩猩一生中至少喪失一個幼崽。實際上,大猩猩幼崽的死因起碼三分之一是由於殺嬰現象。如果雌性只有一個短暫、明顯、公開的發情期,處於支配地位的雄性很容易在此期間獨佔雌性。所有其他雄性因而知道幼崽是它們的競爭對手的後代,於是對於殺死幼崽毫無愧疚之意。
如果雌性擁有隱蔽的排卵期和持久的性的接受能力,她就可以利用這些便利與眾多雄性交配——即便她只能背著配偶偷偷摸摸地做。儘管沒有雄性對自己的父權有完全的信心,許多雄性會認為自己可能就是母親產下的幼崽的生父。如果這樣一頭雄性隨後成功地驅逐了雌性的配偶並接管了她,雄性就不會殺死幼崽,因為它很可能是自己親生的。他甚至會保護或以其他形式幫助幼崽。母親隱蔽的排卵期還將有助於減少種群中成年雄性的爭鬥,因為既然單單一次交配不太可能懷孕,也就不值得為此大動干戈。
作為瞭解雌性如何廣泛運用隱蔽的排卵期的方法之一,可以觀察一下非洲猴中的黑長尾猴,任何曾參觀過東非狩獵公園的人對於這種猴子都不會陌生。黑長尾猴多至7只成年雄性和10只成年雌性群居生活。由於雌性黑長尾猴不顯露任何解剖學上或行為上的排卵特徵,生物學家桑迪·安德爾曼(Sandy Andelman)找到了一棵有一群黑長尾猴居住的刺槐樹,站在樹下用一個漏斗和瓶子收集雌性排泄的尿液,分析尿中激素指標以判斷排卵期。安德爾曼還觀察它們交配,發現雌性在排卵前很久就開始交配,並維持到排卵後很長時間,直到孕期的前半段才達到性接受能力的頂峰。
在孕期的前半階段,雌性的腹部凸出並不明顯,而被欺騙的雄性也覺察不到自己是徹頭徹尾地浪費精力。雌性在孕期的後半階段終止了交配,這時,它已經不能再瞞過雄性了。種群中多數雄性仍有充裕的時間與多數雌性交合。三分之一的雄性可以和每一個單身雌性交配。因此,通過隱蔽的排卵期,雌性黑長尾猴使得近鄰中幾乎所有潛在的殺手保持善意和中立。
簡而言之,赫爾迪認為隱蔽的排卵期是雌性的進化調整,藉此減弱成年雄性對它們幼崽存活的巨大威脅。和亞歷山大·努南認為隱蔽的排卵期可用來分清父權、鞏固一夫一妻制的觀點不同,赫爾迪認為它的功能在於混淆父權,和一夫一妻制無關。
在此問題上,你也許會開始探究“居家父親”理論和“多父”理論都有的潛在問題。因為兩者中的任何一種理論都認為女性有必要對男性隱瞞排卵期,可是為什麼連女性也無法確知自己的排卵期呢?例如,為什麼不能在一個月中的每一天保有同樣紅色的臀部來欺瞞男性,與此同時保有對排卵期的敏銳感覺,並在非排卵期假作興趣高昂來敷衍好色的男人呢?
答案顯而易見:女性在她感到沒有性要求和明知不能受孕時難以假裝出令人信服的性慾。這個答案尤其適用於“居家父親”理論。如果一個女性長期保持一夫一妻關係,配偶雙方彼此熟知,除非她同時自欺否則難以欺人。
無疑“多父”理論對於動物(也許還有傳統人類社會)來講不無道理,在這些物種中殺嬰是一大問題。但這一理論和現代人類社會不甚協調。是的,婚外性行為時有發生,但是對父權的懷疑仍屬個案,而不是推動社會的規則。基因測試顯示至少70%,也許高達95%的美國和英國嬰兒是婚生的,換句話說,是由其母親的丈夫所生。絕少有這種情況:許多男人一邊圍繞在嬰兒周圍,溫情脈脈,甚或送上禮物,提供保護,一邊又在想自己也許是這個嬰兒的生身父親吧。
因此,今天保護嬰兒不受殺戮不可能是推動女性保持長久性慾的原因。不僅如此,我們還將發現,女性也許在遠古時代有過以上動機,但此後性則擁有一種不同的功能,並且保持至今。
那麼,我們該如何評估這兩種相互對抗的理論呢?如同許多其他人類進化的問題,這一問題也無法通過化學家和分子生物學家喜用的試管實驗得到解決。確實,如果世界上存在著這樣一組人群,我們可以使其中的女性在動情期陰部變成鮮紅,而在其他時刻保持性冷淡,使其中的男性只對陰部紅色的女性才興奮,通過這個試驗問題就會迎刃而解。我們就可以看到結果究竟是丈夫花心、無視孩子(如“居家父親”理論所預言),還是排斥異己並殺嬰(如同“多父”理論預言)。可是在科學上,這樣的實驗目前仍不具備條件,並且即便基因工程成為可能,它仍然是不道德的。
但我們仍然可以求助於另一種進化生物學家推崇的強有力的技術手段來解決這些問題,它被稱作比較法。在隱瞞排卵期這方面,我們人類並不是獨一無二的。儘管這種現象在一般哺乳動物中是比較獨特的,可在高等靈長類動物(猴子和猿)中卻相當普遍,我們人類也隸屬於這一哺乳動物群體。很多靈長類動物排卵時並無外部可見的徵象,也有很多雖有徵象,但極細微;還有一些則公然昭示。每一物種的生殖狀況都反映著這麼一種由自然界主持的、有關隱蔽排卵期利弊的實驗的結果。通過比較各類靈長類動物,我們就可以得出哪些是排卵期隱蔽的物種的共性,並發現排卵期公開的物種所不具備的特徵。
這種比較使得性習性的研究柳暗花明。瑞典生物學家比吉塔·西倫-圖爾伯格(Birgitta Sillén-Tullberg)和安德斯·默勒(Anders Møller)的一個重要研究就以它為主題。他們的分析分四個步驟進行。
步驟一:西倫-圖爾伯格和默勒力圖悉數列出所有(共68種)高等靈長類動物排卵期的可見徵象。啊哈!你可能立即會反對,誰有特異功能能洞悉箇中奧秘?一隻猴子發出的信號,例如氣味(信息素)對另一隻猴子是極其明顯的,而人類卻對此無動於衷。比方說,養牛人試圖給一頭良種乳牛人工授精,但卻面臨弄清母牛排卵期的困難,公牛卻可以輕而易舉地從母牛的氣味和行為中識別出來。
儘管我們不能忽視上面這個問題,但高等靈長類動物的問題要比它簡單一些。多數靈長類動物和人類一樣是日出而作,日落而息,非常依賴視覺。一隻嗅覺失靈的雄性恆河猴仍能從輕微變紅的陰部識別出一隻處於排卵期的雌猴來,儘管這種紅色遠不及雌狒狒的來得明顯。而那些我們人類歸類為沒有排卵期可見徵象的猴類,雄猴同樣無計可施,時常可見它們完全不適時地進行交配,例如和未發情的或者懷孕的雌猴交配。因此,我們對於“可見徵象”的判斷結果並非毫無用處。
該分析的步驟一的結果揭示,有將近一半的靈長類研究對像(68種中的32種)同人類一樣缺乏排卵期的可見徵象。這32種動物包括黑長尾猴、狨、蛛猴和猿類之一的猩猩。另有18種動物,包括我們的近親大猩猩,只顯現輕微徵象。餘下18種動物,包括狒狒和人類的近親黑猩猩,則將排卵期公開。
步驟二:接著,西倫-圖爾伯格和默勒按照婚配方式將此68種動物分類,包括狨、長臂猿和許多人類群體的11種物種實行一夫一妻制。23種物種,包括部分人類群體和大猩猩,是由一個成年雄性統領一群雌性,實行多偶制。但是靈長類動物中的多數,即包括黑長尾猴、倭黑猩猩和黑猩猩在內的34種動物則處於亂交狀態,它們中的雌性慣常與眾多雄性交往和婚配。
這時我又聽到某些人在嘟噥,為什麼不將人類自己也歸入那亂交的一類呢?因為我是謹慎地按照習慣進行分類。不錯,多數女性在其一生中前後有多個性伴侶,也有許多女性有時同時和多個男性交往。可是,對一個女性來講,在一個動情週期,她的慣常做法是只與一個男性交往,而雌性黑長尾猴和雌性倭黑猩猩在此期間要和幾個性伴侶發生關係。
步驟三:作為倒數第二個步驟,西倫-圖爾伯格和默勒將步驟一和步驟二綜合起來發問:排卵期明顯程度的強弱是否可能與某種特別的婚配方式有關?根據對前述兩個相互對立的理論的樸素理解,如果“居家父親”理論正確的話,隱蔽排卵期應是一夫一妻物種的特徵;反之如果根據“多父”理論,這一特徵應屬亂交的物種所有。事實上,作為研究對象的一夫一妻的靈長類物種中的絕大多數(11種中的10種)的排卵期是隱蔽的。任何一種一夫一妻的靈長類動物都不會赤裸裸地顯示排卵期,而亂交的物種通常卻會公開顯示(18種中有14種)。看來“居家父親”理論贏得了強有力的支持。
可是,推測和理論之間的一致性僅僅只是事情的一半,我們完全沒有考慮另一半的相關性。儘管多數一夫一妻的物種都擁有隱蔽的排卵期,可是排卵期隱蔽並不能理所當然地推導出一夫一妻制。在32種排卵期隱蔽的物種中,有22種並非單一配偶,而是亂交的或者擁有配偶群。一夫一妻的夜猴、通常是一夫一妻的人類、多偶制的葉猴以及亂交的黑長尾猴都有隱蔽的排卵期。因此,不管什麼因素是導致隱蔽的排卵期演化的第一原因,緊隨其後的一定是這變化多端的婚配方式。
同樣,儘管多數公示排卵期的物種是亂交的,但亂交狀態並不必然推出排卵期的公開性。事實上,多數亂交的靈長類動物(34種的20種)要麼是排卵期隱蔽的,要麼只有很輕微的徵象。擁有配偶群的物種究竟排卵期是隱蔽的、輕微可覺的還是明顯的,也依物種有所不同。這些複雜情況提醒我們,隱蔽的排卵期根據其婚配方式的不同,有著不同的功能。
步驟四:為了區分不同功能的變化,西倫-圖爾伯格和默勒聰明地研究起現存靈長類動物的系統樹來。他們希望據此區分出靈長類動物進化史上排卵信號和婚配方式發生進化的各個階段。潛在的推理是某些現存物種彼此之間有密切聯繫,因此它們很可能來源於不遠的共同祖先,可卻有不同的婚配方式和排卵信號的強度。這就意味著婚配方式或排卵信號在近期發生了進化。
謹舉一例以說明這個推理過程。我們知道人類、黑猩猩和大猩猩有98%的基因完全相同,並且可以追溯到距今最近的約900萬年前的共同祖先(“失落的一環”,即被推定存在於人類和類人猿之間的動物,尚未發現)。可是這共同祖先的三種現代後裔卻呈現出三種排卵徵象:人類的隱蔽排卵期,大猩猩的輕微徵象,黑猩猩公開排卵期。因此,只可能有一種後裔在排卵期徵象方面與“失落的一環”相仿,而另外兩類後裔必定演化出了不同的排卵徵象。
事實上,多數原始靈長類動物現存物種的排卵期都有輕微徵象。因此“失落的一環”應該保持了這一特點,而大猩猩從“失落的一環”身上繼承了這一點(見圖4.1)。在過去的900萬年間,人類卻演進為隱蔽的排卵期,黑猩猩則變為赤裸裸地公開排卵期。人類和黑猩猩在排卵期的徵象方面與我們溫和的遠祖背道而馳。在人類眼中,處於排卵期的黑猩猩和狒狒的臀部腫起很相似。可是黑猩猩和狒狒的遠祖實際上是互不相干地演化出了耀眼的臀部腫起,因為狒狒的先祖和“失落的一環”的祖先在3000萬年前就分道揚鑣了。同樣理由,我們可以歸結出靈長類動物系統樹中排卵徵象發生變化的其他階段。我們發現這種排卵徵象的轉變至少發生了20次之多。明顯徵象的排卵期至少有3個獨立的來源(包括黑猩猩的一例),隱蔽的排卵期則至少有8種(包括人類、猩猩和至少6種不同的猴類);而幾種輕微徵象的排卵期要麼是從隱蔽的排卵期演化而來(如有些吼猴),要麼是從明顯徵象的排卵期變化而來(如許多恆河猴)。

圖4.1 排卵徵象的系統樹
用以上分析排卵徵象的同樣方法,我們還可區分靈長類動物系統樹中婚配方式發生變化的階段。所有的猴和猿類的共同祖先可能都是亂交的。但如果我們審視人類以及人類的近親黑猩猩和大猩猩,你會看到如下三種類型的婚配方式:大猩猩的配偶群,黑猩猩的亂交,人類的一夫一妻或者配偶群方式(見圖4.2)。因此,在900萬年前“失落的一環”的3種後裔中,至少有2種改變了婚配方式。有其他證據顯示“失落的一環”以配偶群方式生活,所以大猩猩和部分人類群體也許是保存了這一婚配方式。但黑猩猩獨自演化為亂交,許多人類群體則是一夫一妻制。我們再次發現,人類和黑猩猩在婚配方式上如同排卵徵象一樣背道而馳了。
總之,情況表明一夫一妻制在高等靈長類動物中至少可分為7組獨立演化:人類、長臂猿和至少5種不同的猴類。配偶群式則至少分為8個分支演化,包括“失落的一環”。黑猩猩和至少2種猴類則在它們的最近祖先賦於它們配偶群方式後自行演化為亂交方式。
因而,沿著靈長類動物的系統樹,我們將始祖靈長類動物可能具有的婚配方式和排卵徵象進行了重新組合。現在我們終於能將兩組信息放在一起考慮:系統樹中隱蔽的排卵期演化的各個階段中,婚配方式又是怎樣的呢?
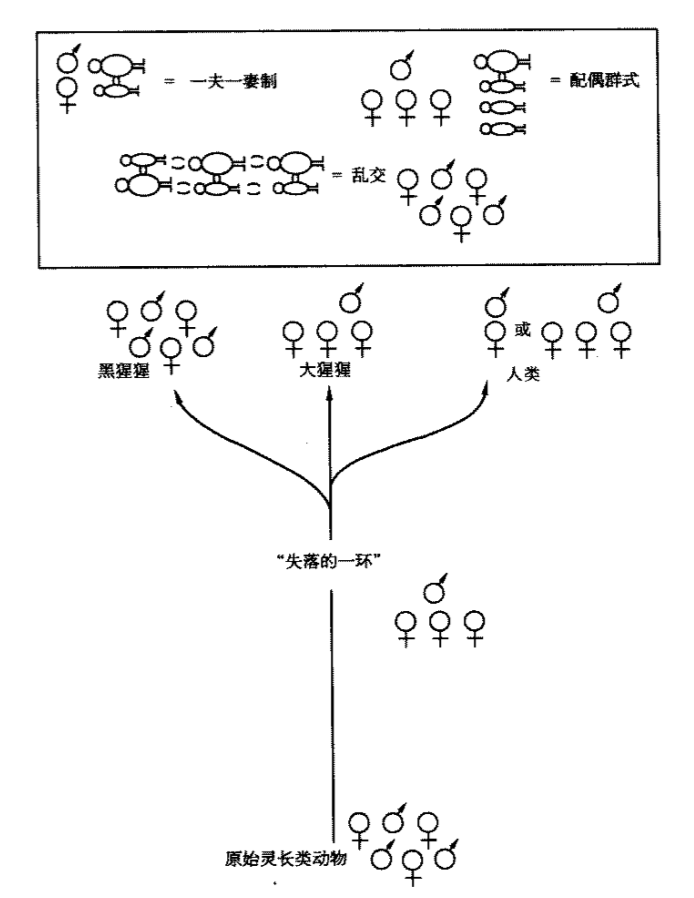
圖4.2 婚配方式的系統樹
下面即是我們的結論。經過考察,那些顯示排卵徵象以及漸次喪失徵象以致演化為隱蔽的排卵期的始祖物種中,只有三種是一夫一妻的。相反,有8種(也許多達11種)是亂交的或者是配偶群式的,其中之一便是起源於以配偶群方式生活的“失落的一環”的人類先祖。我們因此可以斷言是亂交方式或者是配偶群方式,而非一夫一妻制導致了隱蔽排卵期的演化(見圖4.3)。這正是“多父”理論推測的結果,而否決了“居家父親”理論。
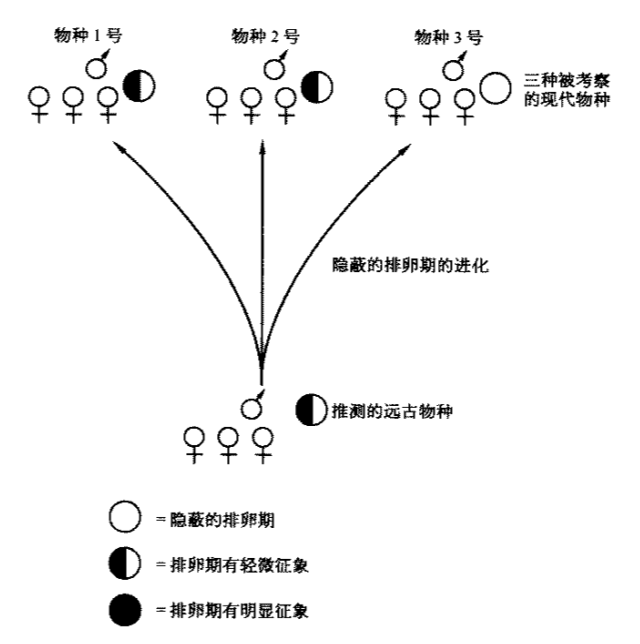
圖4.3 隱蔽排卵期的演化
與之相應,我們又要問:系統樹中一夫一妻制演化的各階段中,排卵徵象又是怎樣的呢?我們發現那些明顯公開排卵期的物種從未採納一夫一妻制。相反,一夫一妻制常見於那些已具備隱蔽排卵期的物種和一些只有輕微排卵徵象的物種(見圖4.4)。這個結論倒和“居家父親”理論契合。
那麼這兩個截然相反的結論是怎樣協調的呢?回想西倫-圖爾伯格和默勒在他們的分析步驟三中的發現,幾乎所有的一夫一妻的靈長類動物都擁有隱蔽的排卵期。我們現在可以發現這個結果分兩步得出。首先,隱蔽的排卵期產生於亂交的或者配偶群式的物種;然後,隨著隱蔽的排卵期出現後,物種向一夫一妻制轉變(見圖4.4)。

圖4.4 隱蔽的排卵期的變化
也許現在你發現人類的性進化史雜亂無章,令人費解。我們開始只提出了一個顯然很簡單的問題:為什麼人類隱瞞排卵期,每月每日追求性樂?照理也應有一個簡單的答案。然而,我告訴你的卻是一個複雜得多的答案,而且包括兩個步驟。
這意味著在靈長類動物的進化史上隱蔽的排卵期的功能一再變化,並且實際上是反向變化著。這種變化起源於我們的祖先仍是亂交的或者配偶群方式生活的時代。在那些時代,隱蔽的排卵期使得始祖女猿人施愛於眾多雄性猿人,沒有一個雄性可以聲稱自己就是她孩子的生父,但是每一個雄性都知道自己可能是。結果是,任一潛藏殺機的雄性都不會傷害這個女猿人的嬰兒,有些雄猿事實上還負起了保護和助養之責。一旦女猿人為此目的演化為隱蔽的排卵期,她就可以利用這一點挑選一個優秀的穴居男性,誘使或者強迫他留在家中陪伴自己,並基於對她孩子的父親身份的確認,為孩子提供許多保護和幫助。
細想起來,我們也無需對隱蔽的排卵期的功能轉化大驚小怪。這些變化在進化生物學上是極為普遍的。這是因為自然選擇不是有意識地沿直線向一個早就認識了的目標前進,也不是按照一個工程師有意設計新產品的途徑進行。恰恰相反,某種動物發揮某種功能的特徵在一開始是提供另一種功能的,當其作為結果出現時已經改頭換面,甚至可能失去了它最初的功能。在生命的進化過程中,相似的功能變化經常再現,經常出現功能丟失、轉變,甚至出現功能的反轉。
人們最耳熟能詳的例子之一是脊椎動物的四肢。始祖魚用來划水的鰭演化為始祖爬行動物、鳥類和哺乳動物的腿,用來在陸上奔跑跳躍。某些始祖哺乳動物和爬行鳥類的前腿,後來又相應演化為蝙蝠和現代鳥類的翼翅,用於飛翔。鳥類的翅膀和哺乳動物的腿又分別獨自演化成企鵝和鯨的鰭狀肢,又回復到划水的功能,有效地再現了魚鰭的作用。至少有3種魚類後裔分別喪失了它們的四肢,變成蛇、無腳蜥蜴和無腳兩棲動物(如蠑螈)。生殖生物學上的特徵,諸如隱蔽的排卵期、明顯公開的排卵期、一夫一妻、配偶群以及亂交,其功能基本亦按同一方式一再變化,彼此轉化、再現或者丟失。
這些進化轉變的意義更增加了我們對人類性愛的好奇心。例如,在德國著名作家托馬斯·曼的最後一部小說《騙子菲力克斯·克虜爾的懺悔錄》中,菲力克斯在一次乘火車旅行時和一個古生物學家同一包廂,古生物學家講述脊椎動物四肢的進化史令他聽得津津有味。菲力克斯這樣一個專事勾引社交界愛幻想女性的男人,竟對此極感興趣,“人類的手腳保持著最原始的陸上動物的骨骼!……真令人毛骨悚然!……女人那勻稱迷人的胳臂,它在我們求歡時是多麼撩人啊!……可它居然和原始鳥類的爪翼、魚的胸鰭無異……下次我再……我肯定會想到這些……想想那漂亮的胳臂,再想像一下那遠古的骨架吧!”
西倫-圖爾伯格和默勒已經闡明了隱蔽的排卵期的進化過程,你可以據此為你的好奇心找到答案。如同菲力克斯·克虜爾通過脊椎動物四肢的進化滿足好奇心一樣。當你安然享有著穩定的一夫一妻關係,在排卵週期中不會受孕的時候追求著性快樂時回想一下,和你的那些因配偶群而憔悴不堪或者是輪換性伴侶亂交的遠祖相比,你是多麼幸福啊!而這種福氣可能正是由那些將你與人類遠祖區別開來的生理特徵賜予的。令人感慨的是,那些可憐的原始人只能在有限的排卵日裡擁有性,把授精當作人類生物需要的全部,倉促了事,由於急於求成而完全享受不到你能從容享有的快感。