
沒有活組織,就不可能有自覺和人類的創造。活而耗散的系統不會有,除非熵沿著它不可逆的通道在時間上前去。
皮考克(Arthur Peacocke)
《上帝與新生物學》
當你念這句話時,電火花在你的大腦中劈啪爆發。熱帶茂密雨林裡,昆蟲總數高漲低降。一個短鼻鱷卵中某點,注定了鱷魚終身穿戴的一套花紋。在每個這樣的生命圖案之中,時間不停地滴答前去。
就像化學界有化學鍾一樣,生物界也有生物鐘。這些鐘的節奏,雖然彼此懸殊,然而對生命都是少不了的。神經細胞,一眨眼之間激發幾千次。另種細胞裡,物質的濃度以數秒長的週期起伏。組成人體心臟的細胞群體,每分鐘跳動70次左右。而組成整個植物或動物的細胞總集,具有內在的、長達數年的成長髮育週期。所有這些節奏歸根到底都是由分子過程、生物化學過程所控制。要瞭解這些過程,我們可以運用與上章我們討論貝魯索夫-扎孛廷斯基(BZ)反應裡時鐘式振蕩時所用的辦法。唯一重要的不同就是我們現在考慮的體系是活體系。
在BZ反應中,自組織表現為化學活動的旋轉螺旋,其中成千上萬的分子齊步組成時間和空間的宏觀結構。在生物界中,相應的組織過程則是個別細胞聚成多細胞有機體。自組織也包含如昆蟲總數的漲落、人體心臟的跳動之類的有序現象。的確,整個的人體都可以看為一個在時間空間自我組織的複雜單元。
因此不足為怪,生物系統具有和BZ反應類似的內部反饋過程。酵素,在被身體製造出來以後,接著又參與隨後的、和它本身製造有關的過程。該酵素可能激勵,也可能壓制細胞的機制。這種非線性過程的結果很難預測,因為當酵素的量在變的同時,製造酵素的規律也在變。但是生命本身就是一個高度非線性過程。載著反饋過程的藍本的基因,同時也負責調製我們身體應該如何讀、如何理解這同一基因。
生物界富有反饋過程,並且和化學界的BZ反應和布魯塞爾振子一樣,這些反饋過程能產生三種不同的自組織形式。時間上的組織是振蕩;空間的組織是圖案;兩者的結合則是活動波浪在空間傳播。關於生命是如何持續這問題,這三種自組織在一起,給我們提供了不少的見解。
這裡必須認清,反饋有本質上不同的兩種:正反饋和負反饋。正反饋增加系統中的產量,一種配料自催化自身的產生就是一個例子。負反饋則降低產量,例如控制暖氣設備的恆溫器。屋子裡溫度一旦低過某個預先定好的溫度,暖氣設備就打開,將室溫提高到所要的水平。該水平一旦達到,設備就關上。然而不可避免地,房屋將會在設備關了以後繼續加溫,結果是使溫度稍微走過頭。同樣地,當室溫下降以後,當設備再開動時,室溫會比預定的「最低點」少許低一點兒。這樣,負反饋結果是建立一個循環,其中溫度慢慢地上升,下降,再上升,再下降……。這本質上是使系統穩定化,此類反饋一般認為在人體中極為重要,例如血壓的控制。純粹正反饋一般認為對人體不這麼重要,因為它會導致不穩定的行為。可是,對於動物總數在時間上的變化,正反饋是很重要的。至於正、負反饋同時存在,這種情況極為平常,身體中白血細胞的製造就是一個例子。
生物學混沌
前面考慮化學鍾時,我們發現有化學混沌。現在我們考慮生命鐘,情況好像是一樣。雖然關於生物混沌的研究仍屬初步,它看上去對某些重要而往往是不良的現象,應該負有責任。混沌似乎發生在體內組織解體,身體處於非正常動力狀態的時候。有人斷言,心臟病暴發時,混沌便嶄露頭角,一如在某些疾病時好時壞的變化之中。另外有人推測,對混沌的瞭解將有利於用腦電活動分析來預報癲癇發作。它甚至是演化少不了的因素,不過,這些論調不宜過分相信。近年來世上無疑地對於混沌的存在有「趕時髦」現象:一方面報刊科學通訊員將它大肆宣傳,另一方面科研者抓住它,用它來贏得名聲與經濟資助。鼓吹混沌的人認為:混沌到處都有。然而,對於許多生物學中的例子,可以說陪審團仍在考慮之中。就如第六章所說,我們應該辨別內稟於系統的決定性混沌和來於外界噪音的隨機變化。區別兩者,理論上有很好的檢驗方法,但是實際應用起來,並非直截了當。這是因為,如果沒有詳細數據,我們依靠不了現有檢定混沌特徵的方法,例如計算自稱為奇異吸引子的分維數。這些吸引子,就是其中資料最齊備的例子,也還沒有人證明過是滿足嚴格數學定義的。
儘管如此,在生物過程每個層次上,從細胞內的事件到細胞與細胞之間的事件,從有機體體內事件到諸有機體之間的事件,似乎多少的確存在有混沌和組織並列。這些事件都是不可逆過程,而不可逆性同時含有混沌和組織的配方,猶如印度舞神濕婆,一手拿著破壞的火,一手敲著創造的鼓。本章將繼續調查研究生命的這兩方面,希望能徹底領會不可逆耗散式系統在自然界的重要性。
由於熵在增大,所有活體系都在耗散。這種體系也都是動態的,因為其中的過程極端地傾向於演化。用術語來說,我們現在是在研究遠離平衡、非線性耗散系統中,時間上不可逆的演化。一個等價的名詞術語就是「動力系統理論」。有些生物學家和數學模擬專家對「耗散」、「遠離平衡」,以至「非線性」這類字眼感到惱火,把它們看成是別有用心的外行強加於現有學科之上、與題無關的商標。這種情緒雖然可以理解,總屬可惜。對生物學的精明見解有些是從跨學科角度取得的。基於不可逆過程的數學性質,我們將要考慮的每個實例都強調時間之箭的重要性,當然,一個理論僅僅應用成功,還不能證明它就是正確的。但我們相信,不可逆性在生物學中的重要性,已積累的證據是如此確鑿,不容忽視。下面的實例,目的就是提出這些證據。
糖鍾
我們身體中的細胞必須具有集中能源的手段。這樣,細胞才可以與時間之箭交戰,而不被時間之箭拖進熱動平衡和死亡——時間之箭最原始的面貌。一群錯綜複雜的化學反應將食物的能量轉化為細緻的生命機制。植物動物只是在持續遠離平衡的狀態之下,才能產生生命必需的生理秩序。它們的細胞需要能量幫助消化,幫助合成生物化學劑。這樣才能產生濃度梯度,肌肉才能收縮,體溫才能保持,等等。
我們的家庭生活靠煤氣或電。身體中的直接燃料叫「腺甘三磷酸」(ATP),是一種富有能量的關鍵生物分子。如要生命繼續下去,ATP必須合成。ATP載能用的是一個高能化學鍵,它像一個壓緊的彈簧,其中牽涉到化學家所謂的磷酸群,由四個氧原子圍繞一個磷原子組成。ATP一旦失掉磷酸群,就退化為「腺甘二磷酸」(ADP)。反過來,ADP也可以經由磷酸化這一過程,進化為ATP。
綠色植物利用日光把A DP(和糖類)合成為ATP,此過程叫光合作用。而動物則用呼吸製造ATP。動物吸取糖類和脂肪,然後在它們細胞中名為粒線體的特殊操作單元,用呼吸從大氣得來足夠的氧氣,燃燒這些材料。燃燒在一串連鎖反應中進行,這些反應彼此策應好像手錶中的齒輪。產生的廢料就和一般燃燒的結果一樣——水和二氧化碳。此種新陳代謝途徑叫呼吸鏈。
然而,並非所有細胞都從陽光或氧氣中取能。有些用呼吸鏈的二等代用品——酵解,使葡萄糖發酵,將葡萄糖分子一截為二,這樣製造的ATP要少得多。巴斯德1861年對酵母進行實驗時,指出這些貧氧過程的效率遠比富氧過程的效率低。正如一位幽默家所說:「這些實驗證明了,沒有空氣,生命不是不行,只是太貴——就像今日紐約一樣。」原始單細胞有機體,諸如也出現在酸乳酪中和食物中的酵母,即使沒有空氣,也能依靠酵解繼續生存。蠔、綠海龜等生物也這樣,它們大部時間生活在水底。甚至在人體中,酵解也起作用,尤其是在血液輸送有限的地方,例如正在進行劇烈活動的肌肉。
在光合作用、呼吸鏈、酵解這三種能源中,生物化學鍾一直滴答前去。我們瞭解最清楚的是酵母中的酵解。酵母是個別獨立生活的細胞,有一種酵母生活在葡萄皮上面,使葡萄汁變成酒的就是它。發酵時產生的不僅是酒精(乙醇),還有別的產物,總數達500種左右,它們之間的比重決定酒的質量。此過程人們熟知,古代文明人都早已知道。根據布魯塞爾自由大學的巴布羅延茲(Agnessa Babloyantz),「有關酵解最早的記載是埃及某座古墓牆上畫的一套解說圖,該墓屬於法老突特摩西斯三世(公元前1505~前1450年)的酒窖管理官」。
釀酒酵母如何把糖轉化成酒精,這項迷惑、陶醉了歷代科學家的研究,是現代生物化學的一個前例。因為此項研究搞得如此深入徹底,我們對生物化學節奏圖案的瞭解,在此情況下達到最高峰。事實上,到了1940年左右,酵解的整個新陳代謝途徑全搞清楚了。1957年,兌森斯(L.N.Duysens)和阿米斯(J.Amesz)首次指出,酵解過程中,能量並不總是平穩地產生,有時能量以一定的節奏振蕩,過程中各種中介物的濃度也同樣地振蕩,其中最重要不過的就是我們的富能老友ATP。
ATP濃度是否在時間上起伏,完全決定於當時周圍糖和ADP的數量。振蕩在生理調製中扮演何種角色,關鍵在此。細胞中如果只有少許A TP(於是較多的ADP),酵解之門就打開,製造所需的ATP分子,細胞或許從其儲備提取澱粉或糖原;如果細胞中ATP很豐富,例如呼吸鏈工作一直很順利,酵解通道就切斷了。此調製過程叫做巴斯德效應,它實際上被單一種酶索所控制,這是一種大而複雜的生物蛋白分子,能加快某些特定的化學反應。
此酶素叫磷酸果糖激酶(簡稱PFK)。經過幾百萬年專門朝這項功能的演化,PFK每當ADP濃度足夠高時就打開,每當ATP濃度夠高時就關上。可是PFK是一種磷酸制劑,它利用AT P把磷酸群掛在糖分子上,從而使ATP轉化為ADP。而ADP本身當然又是激活此酶素的因素,使它更快地工作。這反饋恰恰就是自組織所需要的那種自催化非線性行為。
目前有數種理論模型,目的是描述糖鍾是怎麼走時的。一般說來最成功的是布魯塞爾自由大學的哥爾德貝特(Richard Goldbeter)和勒菲弗1972年提出、隨後由哥爾德貝特及其他合作者所補充的模型。他們把問題中的所有枝節都刪掉,只留下要點。開頭是用12個連鎖非線性微分方程來描述;最後只用兩個這樣的方程,把全力集中在酶素PFK和生物化學能源分子ATP。
如此極端簡化的好處是:描述其中節奏的方程和描述布魯塞爾振子的方程就很相似了。如上所述,它的化學鍾性質可以表示為一個極限環。如果濃度適當,糖鍾裡ATP和ADP的含量便順著一個圈,像跳華爾茲舞一樣,變化週期大致一分鐘,和實驗值符合很好(圖22)。這樣,酵解節奏是生物界耗散結構,時間上自組織圖案的第一個公認的實例。
有節奏的生物化學過程已發現許多,這些過程有的牽涉單獨的酶素,例如自催化劑辣根過氧化物酶和乳酸過氧化物酶,有的牽涉到一群酶索。一如生物鐘,這些振蕩子對細胞內外信號的傳送以及諸如胚胎細胞有的變為腦細胞、有的變為肝細胞的分化過程,都起作用。和前面講過的情況一樣,凡是秩序以可預測的循環形式出現的場合,混沌也就出現,其中振蕩的頻率幅度不停地做不可預測的變化。有規振蕩和混沌振蕩,在植物用以轉化日光為能的光合作用、細胞「發電廠」粒線體的呼吸之中,我們都不能忽視。
細胞分裂的萬能鍾
控制活體基本成長過程——細胞分裂的單獨的鐘,似乎也是自組織在主控。每當我們體內十兆細胞中有一個分裂時,就有新的遺傳物質被製造、被分開、被隔離。細胞分裂調製得好,就得出諸如耳鼻之間的差異,調製不好就是腫瘤。體內細胞以各種各樣的速率進行分裂。成熟的腦細胞根本不分裂。肝細胞每一兩年分裂一次,而腸壁細胞一天就分裂兩次。在細胞分裂過程中,我們再次同時遇到單向式時間和循環式時間。無論從哪一點來看,細胞分裂都是循環,因為剛分裂出來的細胞和其母體完全相似。細胞以一個固定的時間間隔分裂,每繁殖一次,細胞就打一下拍子。可是一系列這樣的循環,產生出來的是線性時間,因為我們的細胞,多數都是命中注定了分裂繁殖若干次以後就要死亡。

圖22 (a)ADP濃度和ATP濃度的週期性振蕩。(b)ADP~ATP相平面中的軌道逐漸演化成一個極限環。
誰都知道,我們體內細胞並非永遠一直分裂繁殖下去。胎兒細胞分裂50次左右以後,就會死亡。一個40來歲人同樣的細胞,分裂40次以後就停止,而80來歲人的細胞,勉強可以分裂到30次。這次數和整個身體老化程度相應,壽命較短的動物,它們細胞分裂較少次數以後就停止了。未老先衰的沃納症患者,他們的細胞死亡以前分裂次數特別少。
對於體內控制細胞分裂的基本時鐘,近來取得了深刻瞭解。我們的身體從一個單細胞,經一系列分裂而成,每次一分為二。該過程嚴防意外,一步連一步,都經由許多因素預先安排好了。有的因素是細胞的內稟因素,例如細胞一般長到某種大小以後才進行分裂。有的因素則受周圍環境的控制,好比說,要看細胞在整個活體中所處的部位。但是,究竟是何種分子使細胞分裂?對於左右這重要關節的遺傳程序,我們能說些什麼呢?
1980年後半期,研究者對這些問題,採取兩種處理方法,從而取得了大步進展。一是科羅拉多州丹佛城的馬勞(James Mallor)領導之下的一個研究小組和法國蒙沛黎誒分子生物學研究中心的皮卡(Andr Picard)、拉貝(Marcel Labb)、多雷(Marcel Dore)領導下的一個小組,認證了觸發細胞分裂的分子。另一個是納耳斯(Paul Nurse)領導的遺傳學小組證明了這個重要的分子,對於從酶素到人基本都是一樣。納爾斯現在工作於牛津大學的皇家癌症研究基金會細胞循環小組。
細胞可以被看做微小的鐘,擺動於兩個狀態之間(圖23)。在一個狀態下,細胞長大,分裂被抑制。在另一個狀態下,雖然繼續長大,細胞卻進行分裂。細胞來回於這兩個狀態之間,一個來回就相當於鐘的一次「滴答」。酵素的基本遺傳程序中這部活鐘的藍本被納爾斯與其同事發現了。他們第一步是把細胞分裂必需的一百多種基因認證出來。每種基因相應於一種對細胞分裂起作用的蛋白。可是光靠這點我們仍不知道分裂的機制。下一步就是要知道哪些蛋白重要。納爾斯體會到,他如果找到一個基因能加強細胞的分裂,那個基因便相當於一個關鍵性的蛋白。他用鐘的比喻來解釋這道理。要鍾走慢一點兒或者要鍾停不是難事,只要拿走某個鈍齒,某個發條,或者其他一堆元件就行了。但只有使鍾走快的操作才能揭露哪個元件是控制速度的關鍵部分。要找出分裂更快的變種很簡單:一個細胞長得越快,它分裂時的體積就越小。納爾斯和他的小組在愛丁堡大學找到了四個這樣的酵母,起名叫「小變種」。其中之一最有意思,含有一個有缺陷的基因,名為cdc2(細胞分裂控制2),納爾斯把它稱作那年的「新聞變種」。它是一大族名為激酶的成員之一的遺傳藍本,這些激酶把別的蛋白打開、關上,辦法是把一個帶負電的磷酸群放在蛋白上面,使它們形狀改變,從而改變它們的催化功能。看上去,細胞準備分裂的時候,此cdc2蛋白激酶打開許多分裂必需的蛋白。

圖23 細胞鍾:細胞成長和分裂的各個階段。
與這項工作相互呼應,當時還有人研究,是什麼因素使未成熟的卵分裂。結果發現,如果把已經在分裂的細胞加在這樣的卵裡,卵就會分裂。令人驚訝的是:所加的細胞可以來自別的生物,海星、人、酵母、「草履蟲」都可以,青蛙、海星未成熟的卵都會被觸發分裂。很明顯地,控制細胞分裂的關鍵分子,從最原始的生物到演化最高的生物,都沒有什麼大不同。1988年,馬勞在丹佛發現了這麼一個分子。納爾斯問他要不要把他淨化過的蛋白和納爾斯自己的酵母蛋白激酶對照一下。果然是同一個東西。這結果納爾斯總括為「有了!成了!」(Eureka)。
牛津小組的李(Melanie Lee)做了一個漂亮的實驗,證明控制人體細胞鐘的也是同一種酵母蛋白。雖然細胞鐘的詳細機制仍屬未知,但是很明顯,管制細胞分裂的遺傳程序,自從地球上有生命開始,億萬年來實質上沒有多大變化:酵母和人有個共同的祖先,10億年前活在地球上,用著和我們今日同樣的辦法控制細胞分裂,這是達爾文學說一個有力的證明。大自然的細胞分裂時鐘,比作手錶也好,座鐘也好,控制機制都是一樣,都是一個分子振蕩子在調節細胞,讓它來回在兩個狀態之間。這「同一種蛋白大家都用」的現象,在自然界屢見不鮮:每當大自然發現了一個有效方法去執行某個緊要任務,她總是盡可能重複使用這同一個機制。這種保守傾向不難理解:一個必不可少的酶索如果變異,形式如果稍有改變,那很可能就活不下去。它就會不再適合自組織所需的、錯綜複雜的生物化學反饋過程。
瞭解細胞循環如何調節,下一步是數學模擬。就如納爾斯所說:「我們現在已經知道了誰在演戲,演的是什麼。現在我們要知道的是:為什麼某個角色在某個時刻上台。」首先,有一些問題必須解決。現有技術很難測量細胞中某個生物化學劑濃度在時間上的變化。分子生物學家在尋找關鍵分子時所用的手段,大半會破壞原有的空間時間組織。按照納爾斯的說法,「我們想瞭解數據,但數據本身已經失掉活細胞動態描述必需的信息。」
細胞鍾更多的詳細情況得到以後,我們差不多就可以斷言,這一個最基本的生命形式的秘密,遲早將會被動力系統學語言說穿。所有必需的分子步驟都得連成一片,然後建立一個適當的非線性動力模型。美國理論生物學家泰森和考夫曼早年(1975)曾在這方面試過,並且自稱得到了極限環的證據,當時極限環概念很時髦,就像今天的混沌一樣。可是更詳細的實驗研究證明了,他們基於一個特殊模型的結論是錯誤的。按照亞利桑那大學溫弗利的意見,這件事給我們的教訓很清楚:這種非線性數學模型的運用,「必須逐次分別加以考慮,時髦概念的預設是用不著的。」
黏菌社交
讓我們現在跨一步到生物界的次一層,不再討論細胞裡面的組織行為,而是討論細胞與細胞之間的組織。像哺乳動物這樣的高級生物,每個含有億萬個細胞,這些細胞在從卵到生殖的過程中,組織成一個極為精巧的結構。在此過程中的任何機制,我們對它的瞭解都不夠給出一個說得過去的數學描述。儘管如此,用非線性動力學可以理解形態起源,這點似乎沒有問題。這是依賴時間之箭的、最令人驚奇的創造過程之一。
一種叫黏菌的奇怪生物,它介於一堆單獨的細胞和一個有機整體之間。它的學名叫Dictyostelium Discoideum(參見黑白插圖),它妙在有時候是多細胞體(10萬左右個細胞),而別的時候,如溫弗利所說,「這些細胞獨立地來來去去,像蟻群中的工蟻一般。它一如蟻群,是一個『超有機體』,一個遺傳一致的整體,由獨立的、不自私的單元,為著集體利益組織在一起。」
黏菌細胞以細菌為食物。周圍食物多的時候,每個單獨細胞都盡情狂吃,以直接分裂方式繁殖。不用說,理想世界不能持久,這社會遲早要缺糧。到這時為止,諸細胞一直都不管彼此的存在,像孤獨流浪人一樣地行動。現在它們「注意」到彼此了。由於某些目前仍不清楚的理由,某些細胞開始活動,成為「領導」,有節奏地發出一種化學劑,名為環腺甘單磷酸(cAMP)。此化學劑cANP在生物界極為普遍,作用是充當鄰近細胞之間的信使。這個要大家團結組織的軍號聲,以每秒數微米(一微米等於一百萬分之一米)的速度傳播。
細胞一接到信號,便開始朝著領導細胞的方向蠕動,那也就是cAMP濃度高的方向。它們同時把信號放大、傳遞,形成反饋,提供非線性,使更多的細胞向領導集合。細胞以脈動波的形式向中心聚集。
只要看一看黏菌就會使我們覺得,這裡的過程和化學鍾裡的大概差不多。書中圖版部分中的照片顯示,細胞群體聚集形成的同心圓波和螺旋波和出現在BZ反應中的螺旋波極為相像。這種波出現在心臟肌肉的節奏跳動之中(關於這點,本章下面還要詳述),也出現在傳染病菌中,乃至漩渦星系裡的恆星形成波。
細胞一旦形成黏膩的一團,便開始分化,形成一個尖頂,從那裡,cAMP不停溢出。整個一團組織成了一個光亮的、多細胞的「鼻涕蟲」,有一個頭,一個尾,向前蠕動,找光,找水。這群細胞形成這簡單有機體,前後一共要數小時。它身長一二毫米,在脈動源尖頭的領導下向前爬行。過後它豎立起來,形成一個桿子,桿子頂上托著含有孢子的囊袋;最後囊袋破裂,風把孢子吹往遠處。如果孢子停在適當的地方,它們便會發芽,重新開始這奇怪生物的另一次循環。
這種行為裡面令人注目的生物化學,使我們聯想到上面描述糖鍾裡的酵解反應。為此蠕動物體提供信號的信使分子,cAMP,是從ATP經過一種名為腺甘酸環化酶的酶素的作用而產生的。就像酵解一樣,這裡也有反饋出現:細胞周圍介質中已有的cAMP又激發腺甘酸環化酶,從ATP中產生更多的cAMP。這樣便出現了自催化,而自催化是自組織不可缺少的因素。哥爾德貝特運用了他先前研究酵母細胞中酵解振蕩時用的非線性分析,在極限環基礎上詳細地指出,cAMP如何每幾分鐘產生振蕩。這是自組織行為的一個極好例子;再者,現在也有了cAMP混沌振蕩實例。在D.Discoideum的一個變種裡,已觀察到以cAMP振蕩形式出現的時間上的混沌,以畸變枝幹和結果體形式出現的空間的混亂,這些現象,如添加一種名為磷酸二酯酶的酶素就都可以恢復正常。
生物形態的起源
大自然具有多種機制,使細胞組織成為各種各樣、令人眼花繚亂的形狀。如用最簡單的方法分類——按照體積的大小,人的位置便是在離最大頂端的十萬分之一的地方。鳥、鼠、蜂或許看上去不大,其實它們也處於離頂端的1%之處。其餘99%都是昆蟲和小蟲,它們的平均長度僅僅3毫米——「這樣大小的東西,就是掉在湯裡,我們也很少注意到」。要詳細描述多樣化配方,為期尚遠。然而,人在相當大的程度上,受著和其他生物,諸如昆蟲、蒲公英、黑猩猩同樣遺傳程序的控制,這一事實使我們繼續有希望能對演變有個一般性的理解。
有些科學家只想對實際過程略加澄清。有的卻想發現生物界的牛頓定律。20世紀早期,蘇格蘭聖安德魯大學動物學教授達西·湯普遜(D'Arcy Thompson)在他關於多細胞有機體成長的專著中寫道,有生命物體的進化必須受幾何所規定的框架的限制。今日的生物學家,許多都會認為這是條死胡同,認為該學科的大步進展不是出諸筆墨,而是來自實驗室中的觀察。
儘管如此,在生物學實驗研究激增的同時,理論學者也在背後悄悄地致力於瞭解這些生命圖案如何在時間上編織出來。從理論上我們不僅可以得到酵解和細胞鍾所表現的時間上的圖案,我們也可以同樣取得諸如條紋、點子、山形紋等的空間圖案,這些可以看做為駐波:雖然組成花案的彩色分子本身可能在做劇烈活動,整個圖案是固定不動的。此種可能性早被杜靈在他1952年的一篇論文中認識到。關於形態起源——有機形式的發展,杜靈的看法近來有人研究是否可以用來解釋諸如斑馬的條紋、金錢豹的點子以及身體各部分的初始分化。
要使一個圖案從億萬個運動不已的分子混合體中出現,其所需要的配料在本書第六章討論過。其一是擴散;化學渾湯不同的部分到底是經過擴散彼此才「交換信息」。另一個是化學反應。分子一面擴散一面起反應,反饋出現,於是花樣產生。為了對這些空間結構做數學模擬,非線性動力學必須描述不可逆化學反應的速度,同時要顧及各種化學配料不同的擴散率。因此,杜靈的基本途徑現今稱為「反應-擴散理論」。第六章提到的「可激發性」可能也很重要。
生活在淡水裡的水螅,有人說有100個,有人說有50個,有人說有9個頭。雖然古希臘的大力神,借助於其馭者,終於結果了它的性命,它的一個低賤的遠房親戚現今仍然健在,並且在對形態發生的研究中做著貢獻。從水螅身上截取一二毫米的一小段就可以長出整個的水螅。這驚人現象早在1744年就被出生於日內瓦的動物學家特蘭布利(Abraham Trembley)注意到。它為形態起源的研究,提供了一個雖非典型、確是方便的系統。如果從水螅頭附近取出一小片組織,放在身體的另一個部分,48小時以內就會長出一個新頭;那個新頭甚至可以拿走,它自己又會再長。「局部組織受到某個物體的感染了」,馬普病毒研究所的馬因哈特(Hans Meinhgrdt)這樣寫道。馬因哈特對此形成圖案的能力,在杜靈式反應-擴散理論的基礎上,進行模擬。按他所說:「很有可能,在不久的將來,關於簡單有機體成長的控制,水螅將會提供一幅頗為完整的圖畫。」
水螅中圖案的形成似乎取決於兩個因素:短程的化學激發(經由自催化)和遠程的抑制。產生出的非線性導致許多有機體所共有的圖案特點。典型情況是:一小片組織開始與其周圍稍有不同,開始釋放少量的「激發劑」,由於自催化,激發劑的濃度很快地增高。局部的高濃度觸發抑制信號的製造,這是另一種生物分子,它散佈到周圍的組織,使別處不生產激發劑。這些所謂「形態子」的濃度分佈輪廓,實際上告訴細胞它相對於特殊組織的位置,從而決定此細胞演化為頭細胞還是尾細胞。例如,要看是昆蟲身體的哪一段,結果是腿還是觸角。一般認為,激發和抑制不僅模擬初始圖案的發展,並且對諸如鬃毛、毛髮、羽毛、樹葉重複式結構的間隔,也扮演重要角色。反應-擴散理論已被運用於四肢軟骨的排列,羽毛鱗甲的分佈,動物的體紋以及蝴蝶翅膀上的複雜花紋。是否成功,尚有爭論。反應-擴散式方程可以產生各式各樣不同的圖案。杜靈的母親寫道:「他給我看了一些(圖案),問我它們像不像牛身上的斑點。它們的確很像,以至於現在我一看到牛就想起他的數學圖案。」
動物的這種圖案一般規定在殼或子宮裡的胚胎上面。圖案形成的精確時刻和當時胚胎的大小,是決定動物終身穿戴的花紋的關鍵因素。數學模型啟示我們,為什麼處於哺乳動物體積分佈譜兩端的象和老鼠,它們身上的顏色比較均勻一致,而不太大也不太小,像短鼻鱷魚、金錢豹、斑馬,它們身上的花紋就會很不尋常。此類模型表明,金錢豹的尾巴太細,裝不了斑點,斑點都合併為條紋。的確,按照牛津大學數學生物學中心的詹姆士·默裡(James Murray)教授所說,數學模擬可以解釋為什麼「世上有身上是斑點、尾巴是條紋的動物,而沒有這個樣子反過來的動物」。
然而,用杜靈不穩定性來描述圖案形成有個大問題,那就是:該不穩定性從來還沒有在任何實驗中真正出現過。對有些似乎控制發育的物質,有人建議它們是「形態子」:哈佛醫學院的一對夫婦小組得到很好的證據,證明視黃醛酸會觸發雞胎中的一團細胞,使它們成長為腿或者翅膀;而坐落在曲賓根的馬普生物演化學研究所的一個小組得到證據,證明果蠅胚胎的發育受bicoid蛋白梯度的影響。可是這類例子不符合杜靈關於花樣產生的圖像。其理由本書第六章已經略為提及。杜靈的反應-擴散理論無法描述自組織所有可能的機制。加州伯克利的吉姆·默裡(Jim Murray)、奧斯特(George Oster)和其他人提出了一個與此有關但性質不同的處理方法。他們用一個類似的模型,但裡面採用直接測量的生化量和細胞密度,他們叫這「力學一化學」方法,因為細胞受到的化學和力學影響都考慮到了。此模型可以和已知量或實驗可測量掛鉤,它是否正確因而比較容易檢驗。默裡使用了一個力學-化學模型,對短鼻鱷魚身上花紋是在懷孕期哪個時刻規定在胚胎上,做了精確的預言,這個具體結果可以直接檢驗。此種措施進一步的運用將幫助我們瞭解,創傷如何自我治癒,正常發育期中四肢的軟骨佈局如何規劃以及是何種因素導致先天性缺陷和畸形。

圖24 在胚胎時期決定的動物身上的花紋和動物的大小有關。老鼠體積大小,因此多數身上沒有斑點。體積大的動物,好比象。也傾向於全身一個顏色。用數學我們可以預言,面積不夠大時,斑點就要變為條紋。獵豹下方的楔形圖說明了獵豹身上和尾巴上花紋的不同。
跳動的心臟
形態起源是不可逆演化的一個極好例子,但它對我們心理起的作用遠不及那台最了不起的生物鐘——心臟。人的心臟一分鐘跳動70次左右,一年4000萬次,一生總共30億次上下。這台充滿活力的發動機,它所有的各種規則的節拍,都來自於不可逆過程。心臟不僅是生命的象徵,它同時也是生命的一個共同極限:所有哺乳動物小如老鼠,大如海鯨,一生中心跳總數似乎都差不多,約20億次。
心臟演化的前身是比它簡單得多的生物泵。原始動物用這種生物泵,以蠕動的形式,使液體周流其身——今日人體中的大、小腸也是以蠕動形式運輸食物。兩個這樣的管子經過演化彼此盤繞,長出極大的肌肉,最後形成我們今天四個心室的心臟。自從人類在地球上定居以來,有關心臟跳動的作品,從原始部落的歌唱,到歷代的詩詞,直到今日的流行歌曲,屢出不窮。現在輪到數學家和科學家顯身手了。
在20世紀20年代,荷蘭的兩位學者范德泊(B.van der Pol)和范德馬克(I.van der Mark)在這方面種下了種子,直到現在仍在給我們寶貴的啟發。他們指出,只要調節模型中某些參數,便可以得到心臟規則節奏的各種不同的破壞——各式各樣的心律不齊;每種心律不齊的出現都是經由非線性動力學中很熟悉的一個特點——分叉點。
何傑金(Sir Alan Lloyd Hodgkin)和赫克斯利(Sir Jul-ian Huxley)1952年的先驅工作,為生理系統的數學模擬帶來了一大進步。他們研究的是魷魚的巨軸突——神經細胞傳遞衝動的線狀伸延。這工作使他們獲得諾貝爾獎。他們處理所得到的微分方程的方法,能給出一個定量的描述,現在經常被用來研究模擬心臟組織中電活動方程的性質。赫克斯利隨後在1959年又指出,內稟於模型的是重複性活動,如果細胞缺鈣,便會出現規則性的神經激發。
關於心臟動作的情況以及用於描述的非線性方程的數學性質,我們現在知道的比以前多多了,對一大批心臟病的醫療,也取得了驚人的、不斷的改善。例如,我們現在在心臟上可以置放一個器具,監視心臟的跳動,一出現可能致命的節奏,便搖它一下,使節奏恢復正常。很可能的是:不可逆非線性動力學這理論工具,對心律失調的診斷和治療,遲早會有貢獻。僅在美國,一年就有40萬人由於心搏突然失調而死。這裡節奏失調有時是跳動過慢,多半是心搏過速。雖然所謂過速會是快而有節奏的拍子,可是心電圖會揭示,當心臟開始做纖維性顫動時,這拍子就退化為無規則的形式。這表示心臟細胞的一個非正常的時間空間組織。
許多生理節奏是由單個細胞或數個耦合在一起的細胞產生的。就心臟而言,我們現在知道心搏至少受到六種不同的指揮。一個特殊的肌肉組織,名叫普爾金耶纖維,它的細胞比心臟別的組織的細胞都大。雖然普爾金耶細胞並不直接提供心臟的自然節奏,它們把「竇結心搏引導」所發出的激發傳運給心臟。這些引導細胞的節奏是由所謂「心搏波」觸發,它們類似於BZ反應中的引導中心,那些中心可以用熱鉑絲稍碰溶液而建立,一粒灰塵或者盛溶液的碟子上一道劃痕都可以觸發它們。反過來,心臟病發作時也觀測到與BZ反應中類似的螺旋波,圍繞著一塊健康的肌肉組織旋轉。
牛津大學生理系的計算機裡面跳動著一個活的心臟細胞——基於一個數學模型的數字式心臟細胞。這是諾布勒(Denis Noble)和他的小組設計的。諾布勒、弗郎西斯科(Dario di Francesco)和他們的合作者致力於建立一個數學模型,其中包括發生在細胞裡的成千上萬種化學作用。關於心臟跳動的原因,世界上的科學家一發現什麼,他們就把它加進去。可是即使模擬一個細胞的動作也需要大量的計算機時間——要算一秒鐘的心跳,諾布勒的計算機要運行100秒。因此要瞭解眾多細胞的合作行為,會大大受到計算機時間的限制。諾布勒希望用明尼蘇達大學的「連接機」(Connection Machine)來模擬高達50萬個細胞的心臟組織。這是台威力巨大的計算機,具有1萬個可以平行計算的處理器。這類巨型數字運算可能幫助我們對不同的心搏分析加以比較,對一些問題不同的說法加以判斷。例如,纖維性顫動有人說是起因於決定性混沌,有人說原因是旋轉波的相空間奇點。
在本書寫作的時候,細胞中的物理化學過程是用30多個聯立耦合非線性微分方程來描述。這些過程中最重要的是讓電信號閃進閃出細胞的渠道。這些信號由特殊蛋白傳達,辦法是把載電的離子搬來搬去。諾布勒的模型包括十多個這樣的渠道以及另外一些在細胞內部和表面運輸化學劑的過程。這裡面最重要的是鈣渠道,它觸發ATP經過一串複雜的過程轉化為心搏。與BZ反應一樣,心臟也是一個耗散系統,因為物質進出細胞,要使細胞兩邊化學劑的梯度不斷減低。
化學鍾裡面的耗散式結構是規則性的顏色變化或者美麗的彩色漩渦。心臟細胞裡,眾多蛋白纖維配合一致的動作可以看為其中的耗散式結構。為了維持這節拍,個別像鈉泵這樣的過程是不可缺少的。就像座鐘鐘錘漸漸地下落使鍾不停,這種泵不停將鈉離子輸送過細胞膜,從而保證心臟遠離平衡態。每次心跳,鈣離子就湧入心房。跟鈣渠道相互作用的不是別的,而是我們的老朋友cAMP。一如黏菌情況,cAMP和酶素腺甘酸環化酶並肩工作。在心臟裡,這兩位在控制鈣渠道開關的反饋過程中,齊步動作。在單個細胞的層次上,湧入細胞的鈣離子,運用一種分子「棘齒」機制,觸發某種特殊蛋白的收縮。顯而易見,鈣進入心臟只是一半心搏。一次心搏完成以前,必須有另外一個運輸機制,把鈣取出來,放鬆細胞。這現象已經用特製的鈣敏染料看到了。
得到的圖像是依靠一群錯綜複雜的、自我組織的、不可逆反的相互作用的一個心臟——一群化學,蛋白、酶素信使的集體舞組成的每次心搏。諾布勒的模型,以其逼真的跳動,顯示了心臟舞蹈可以用非線性微分方程加以數學描述:是一個極有考究的物理-化學「時鐘反應」使我們心臟搏動。
動力性疾病
諾布勒的研究除告訴我們關於生物時間的一些情況外,還給心臟病的研究帶來了好處。即使正常人,心跳率也有顯著的起伏;當心臟鍾裡有多個彼此競爭的引導中心時,不正常的心搏常會出現,這情況有時就模糊地被說是「混沌性」。現在既然有了一個非線性動力心搏模型,我們就可以考查這種不規則性是來自決定性混沌,還是來自較傳統的原因——隨機噪音。要把兩者從記錄在心電圖上的實驗數據中分開,殊非易事。因此目前對於如何理解此類數據,意見不一。有些作者說混沌將會揭露心臟病暴發的隱秘,但這看法不一定恰當。例如,心室纖維性顫動是一種導致暴死的心搏失調,雖然臨診醫師常說它不規則(意即「難以描述」),可是仍然看不出術語意義中的混沌[圖25(a)]。
蒙特利爾麥吉耳大學的格拉斯(Leon Glass)、圭瓦拉(Michael Guevara)和希瑞爾(Alvin Shrier)對雞胎心臟細胞進行的研究,可能是臟腑層次上生理混沌的資料最齊全的證據。心房細胞一些群體自發地、有規則地跳動著,給它們來個電震,下次的搏動便會提前或者推後。如果電震是週期性的,雞心細胞便受兩個頻率的推動——細胞的內稟頻率和外力的頻率。這是能導致決定性混沌的經典情況。上章注意到,BZ反應中,改變化學配料增加的份量——相當於此處的電震,也同樣導致外表上隨機的行為。在酵解實驗中,介質中糖的流動如有週期性變化,同樣的現象也已觀察到。類似地,取決於電震的頻率,雞心細胞在兩次搏動之間的激發,也許是數目一定的,也許是混沌式的。
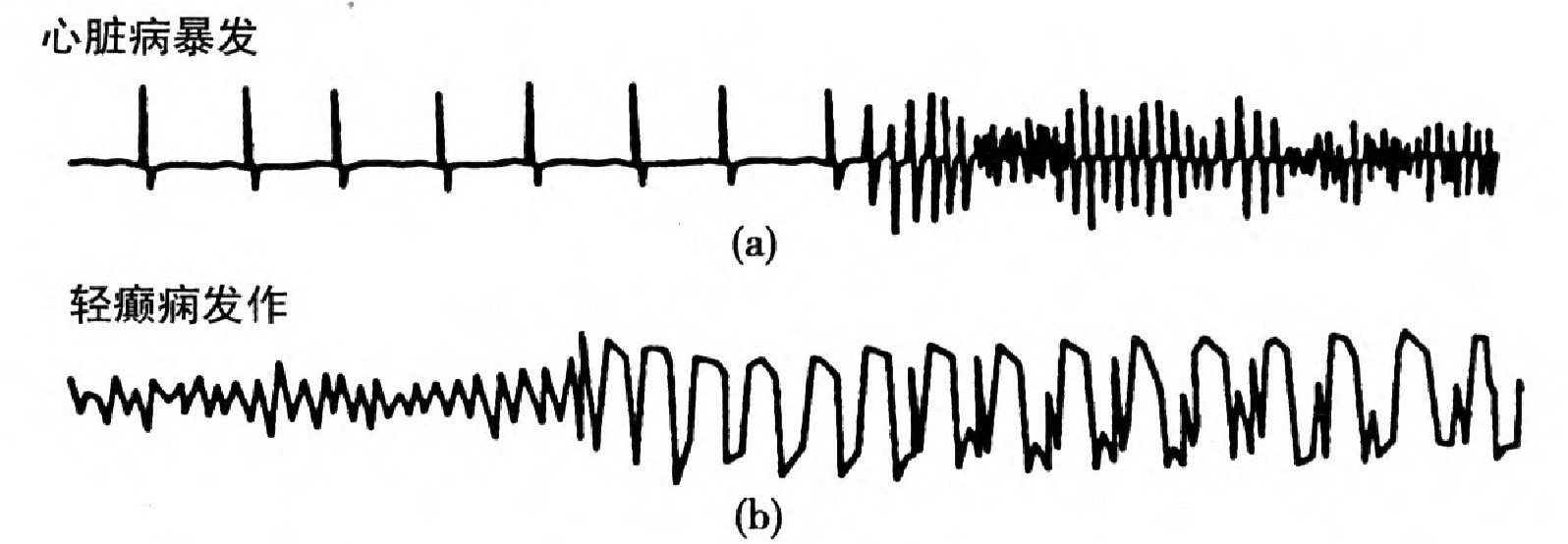
圖25 兩種情況之下的節奏變化。(a)心臟病暴發;(b)輕癲癇發作。[錄自美國《科學》第243卷,第604頁(1989)。]
批評者也許會說,雞細胞的實驗研究人為性很大;即使這裡出現的是決定性混沌,它也不一定會自然地出現。波士頓的貝絲·以色列醫院的哥爾德貝爾格(Ary Goldberger)卻認為會。哥爾德貝爾格在心電活動分析的基礎上甚至宣稱:健康人的心臟比病人的更為混沌。這表面奇怪的結論似乎違背常識,我們總以為許多身心功能的失調是由於健康機體節奏的瓦解。哥爾德貝爾格反而認為,混沌屬於健康身體,而疾病跟此靈活性的喪失有關。對此觀點,頗有爭論。
溫弗利採取了另一種立場。1987年,他猜想心臟肌肉如果在適當時刻受到適當大小的刺激,便會有出現旋轉波式的心搏失調的危險。此後在心臟中誘發這種很快導致纖維性顫動的旋波的實驗,支持了他這猜想。實驗發現,如果電擊施於心臟循環中某些特殊相位,順、逆時針方向的旋轉波都可以觀測到。據溫弗利說:「生物醫學受物理理論引導的例子,少得可憐;這個實例真太令人滿意了。」他對我們目前的理解,仍抱樂觀態度。溫弗利在他的書《時間破碎時》結論部分寫道:「我首次遇到纖維性顫動這詞是七年前,現在我還是不知道它的含義……它仍是謎。應該有人解這個謎了。」
心臟病暴發時規則性心搏的破壞,是格拉斯和麥齊(Mi-chael Mackey)所謂「動力性疾病」的一個例子。動力性疾病是由於身體正常節奏的移動而產生的。這定義巧妙地避免了混沌是好東西還是壞東西的爭論;只說病是因為節奏變了。這種動力性疾病,醫生們都很熟悉,實例包括癲癇發作和各種呼吸失調,例如「澈恩-司托克斯呼吸」(Cheyne-Stokes breath-ing)——呼吸週期性地變快變慢,往往伴隨心臟充血衰竭。不同的疾病像心臟病發作和癲癇,現在可以作為動力性疾病的例子進行比較。這從簡稱為EEG的腦電圖[圖25(b)]可以明顯看出,腦電圖顯示的是活人腦中的電波,這些電波的來源是十萬分之一伏特的電干擾。
腦電圖是由放置在頭皮上的電極記錄得到;這是每個醫院的常規工作。電極和狀如蕈傘的大腦外皮調準。和心搏不同,一個正常健康人的腦電圖是不規則的,並且是比較寧靜的。當癲癇發作時,此活動起劇烈變化,此時病人會昏迷過去不省人事。中古時期,癲癇發作被認為是惡鬼附身,或者就是魔鬼本人。可是並不表現為腦神經失常。因為這時的腦電圖中的電活動,雖然幅度比較大,節奏反而更有規則些。
巴布羅炎茲(Agnessa Babloyantz)與其布魯塞爾自由大學的合作者,對癲癇發作時記錄的腦電圖,做了詳細的分析。腦電圖究竟能提供多少信息,不太清楚,因為用以記錄的電極測量的是大腦頗大區域的平均電活動。儘管如此,這群科學家運用了耗散非線性動力學分析的標準技術,宣稱在正常人和癲癇患者的腦中都找到了奇異吸引子——混沌的標誌。這數學抽像的分維數大致表達混沌的程度:分維數越大,隨機性也就越大。從腦電圖的數據,他們計算得到的分維數是有變化的。對於一個活躍的正常頭腦,他們未能把分維數算出來,他們只知道這時的分維數比熟睡時要高得多,而熟睡時的分維數少許高過4。但癲癇發作時,腦活動變為更有秩序,分維數幾乎下降到2。
耗散式動力系統理論對體內節奏所提供的新認識,很可能加強目前醫術的效果。一個引人入勝的課題是慢性骨髓白血病患者的生存率,這生存率50年來沒有改進。有人認為,這是因為醫生們沒有充分注意到白細胞數本身也是有漲落的。格拉斯和麥齊相信,進一步瞭解控制這些節奏的系統,將會導致更有效的醫療。
對於病人經受刺激以後有時出現的各種意外,動力學見解或許也會有幫助。這裡所謂刺激可以是施諸心臟的電震失控,也可以是來自機械式血液淨化器。最平常的情況是阿司匹林的經常服用或某種針藥的經常注射時,進入人體的某種藥劑。在此情況下,在身體自然節奏和治療外加節奏之間,很不容易建立起穩定的關係。這告訴我們,應該尋找新途徑來改善現有藥品的服用,對此,有些藥材公司已經開始考慮。在不久的將來,對生理節奏的理解會大大改進,醫生們將能利用這一成果。有人甚至說,找出正確的服藥時間,可以抑制規律性癲癇的發作,把刺激時間調好,可以抑制發抖,等等。
科學家們現在正在從非線性動力學中取得諸如此類的認識。誰想到過,從抽像理論的研究中,出現了醫學上重要的進步?手裡控制科研經費的政府官員、官僚、政客,他們大多數肯定沒有這樣想過。
性循環或硬激發
甚至性別,也被非線性動力學所表示的時間之箭控制。第五章給的分叉圖理想化地顯示出,當系統被驅趕到遠離平衡態以後,所出現的可能性。在化學鐘的情況下,出現了多種狀態,這實際上意味著從一個具有規則顏色變化的振蕩狀態轉變為一個非振蕩狀態。過渡方式之一,名叫「硬激發」,也叫做「次臨界霍普夫分叉」。這意思是:當某個參數(例如某反應劑的濃度)增大時,以前一直沒有的振蕩或節奏突然發生。相信性交高潮就是生理方面的一個例子。
一個健康男性射精時,他骨盆底部肌肉的電活動如果記錄下來,便可看到活動是爆發式的——是一串速射的神經衝動。此種事前沒有顯著的週期而突然發生的爆發,是和「硬激發」一致的,這點,讀者或許會感興趣,然而不會驚訝。已經發現的還有其他這種節奏突發的例子,包括婦女停經期陣發性發熱感。這種現象的反面也同樣會發生,原有的振蕩突然地消失。有人認為這就是嬰兒忽然停止呼吸猝死的機制。
群體動態學
我們現在把注意力從生物體內的事件轉向生物之間的事件。描述這些事件的數學,也同樣具有時間之箭。這裡一個很好的例子就是群體生長率,早在1220年,比薩城的雷歐納多[又名菲波納其(Fibonacci)]對此做了數學模擬。他給出一個極為可怕的預言,說一對兔子,如果讓它們繁殖114代以後,它們子孫的體積將大過所知道的宇宙。隨後有人指出,「遠在此以前,地球就被埋在以超光速膨脹的兔子球體裡面了。」
由於存在有掠奪、疾病、競爭、合作,大自然不會那麼多產,那麼不靈巧。兔子群體和吃兔子的狐狸群體,在同一個生態系統中進行錯綜複雜的演化。地球上的生命,競爭演化,彼此糾纏的程度,不亞於細胞中各種分子組成的交響樂。有人把細菌、哺乳動物、植物、魚看做是一部周轉日光、養料,全球性活機器的個別齒輪。19世紀,昆蟲學家記錄了寄生蟲與其宿主之間有節奏的變化,開始意識到這種相互依存。大自然中的均衡必須調節得很準,否則某種生物會把它捕食的生物吃盡,然後自身餓死而絕種。
20世紀初,人們開始用數學模型來描述群體在時間上的變化。這類模型不可避免地含有時間之箭。結果得到是耦合的非線性微分方程,顯示著比方說狐狸總數和兔子總數之間的相互依賴和它們各自在時間上的演化。群體動態學中最重要的成分是競爭(其實這也是演化的最重要成分,演化下面接著就講)。兔子數目不太小時,兔子是狐狸的犧牲品,狐狸總數增加,兔子總數減少。可是一旦兔子太少,狐狸總數就要降低。這樣一來,兔子總數將重新增高,整個循環又重複一次。這種對有限資源的競爭造成一個起調節作用的反饋機制,該機制從數學觀點來看,又是來自某些非線性。這相當於化學鍾反應裡的自催化。美國的勞特卡(Alfred Lotka)提出第一個振蕩式的掠奪-犧牲模型,該模型由伏爾泰拉(Vito Volterra)獨立地加以充實,伏爾泰拉是第二次世界大戰前意大利科學界有影響的人物。此模型現在大家叫勞特卡-伏爾泰拉模型。
然而,這種時鐘式的行為,這種規則性的起伏,在動物界中並不常見。這方面最詳細的長期記錄大概要算加拿大山貓(猞猁猻)總數的記錄,原因是200多年來人們一直為其皮革獵捕山貓。從這些記錄可以看到山貓總數有大幅度的起伏(圖26)。這複雜的變化,許多人猜想是反映著山貓吃的動物——雪鞋野兔的總數的漲落,而這漲落又來自野兔食物數量的變化。可是勞特卡-伏爾泰拉模型不能用來描述這批數據,如果硬拿來用,就會得出野兔吃山貓的結論。
借助於非線性動力學,有人提出了另一種解釋,其中只牽涉到山貓和野兔,而不管野兔食物的供給、天氣的變化、疾病或其他外界因素。一個非線性動力學系統裡。不規則起伏可能來自混沌。事實上,生物群數理論學者把決定性混沌這概念放在舞台中心,就是因為群數無規律的漲落是很普遍的、迫切需要解答的現象。「混沌」這詞就是一位理論學者,馬裡蘭大學的約克(J im Yor ke)在1974年創造的(研究報告發表於1975年)。
這種混沌現象的數學性質,梅爾堡(Myrberg)早在1962

圖26 加拿大山貓群數在1820~1930期間有顯著的漲幅,每九年、十年達到高峰隨著是迅速的下降。
年列出,隨後又被其他人獨立發現。梅(Robert May)無意中碰到非線性系統這些多得令人迷惑的可能性,1974年在美國的頭號雜誌「科學」上撰文,從而成為把它介紹給生物群體學者最早的作者之一。「現在回顧起來,很奇怪這種混沌動力學沒有人更早注意到」,梅說。梅現在工作於牛津大學。在非線性動力學道路上,梅在試圖大步行走之前,是緩慢地匍匐而行。他首先研究可能是最簡單的群體動態模型——非線性「運籌方程」,此模型在母女不並存的情況下,對物種總數的變化,給出一個簡要的描述。這裡一個好例子是春天蜉出、秋天產卵後就死去的昆蟲群體。這群體由於出生率的大小和對食物的競爭,會有多種不同的演化方式。出生率和死亡率可能相等,那時群體就處於一個定常狀態,就不再變化。用動力系統理論的語言來說,這就屬於一個定點吸引子。另一種情況是:群體總數可能出現有規則的跳動,在一些不同的定點吸引子之間跳來跳去,出現於兩個、四個或其他數目的固定值之間。再有,總數可能似無規律地上下起伏,那時就相當於奇異吸引子所描述的混沌(圖27)。

圖27 同一模型(運籌圖)中的各種行為;(a)穩恆狀態;(b)2週期;(c)4週期;(d)混沌。[錄自美國《科學》第243卷,311頁(1989)。]
整個行為譜包含在單一方程之中。每種行為,只要換一下輸入方程的一個參數,就可以算出來。變動此參數而得到的這假設的昆蟲群體所有可能的選擇,可以用我們熟悉的分叉圖來說明。用計算機圖解,可以把所有可能性畫為一大幅奇怪的圖案,圖案的每一點代表一次計算的結果(參見彩色插圖)。這些圖是德國多特蒙德市馬普營養生理學研究所的馬爾庫斯(Mario Markus)與海斯(Benno Hess)繪製的,每幅是數萬次計算的結果,考驗非線性方程的全部本領,結果就是這樣。把方程中的參數稍變一點,得到的就是完全不同的另一景象。
梅的工作使人們驚奇:它說明,複雜性是一個非常簡單的方程中的內稟屬性。梅和他的同事奧斯特爾(Geroge Oster)進一步研究運籌方程所屬的方程族中別的成員的分叉圖性質,——所謂「二次圖」的性質。其中有些與生物學似乎較有關連,在他們得到的分叉圖的「樹」中,他們在逐漸減小的尺度上發現有自相似性,從而揭示出一個奇異吸引子的分維幾何。
這項工作的重要性——說明簡單模型會有出人意料之外的結果,是梅在寫一篇評論時忽然意識到的。梅講道:「一旦有了二次圖,道理連初中學生也能懂。只要在一個計算器反覆運算。一個十二歲的孩子也會做。他能親眼看到,一個簡單、確定的規則會做出稀奇古怪的事。」
在刊登在英國《自然》雜誌上的一篇極有影響的評論中,梅指出,群體動態學者所用的數學模型也出現在許多別的學科。除生物學中包括遺傳學等不同的科目外,同樣的方程也出現在經濟學,那裡它們被用來描述商業盛衰週期、商品供應與價格之間的關係,等等。在社會科學中,這些方程被用來描述謠言的傳播。我們可以期望得到這些系統同樣多種類的行為,從規律性振蕩到時間上的混沌。梅的結論說:「不僅科研,即使在政治經濟每日世界中,最好能有更多的人體會到,簡單的非線性系統並不一定具有簡單的動態性質。」如果主宰經濟的是決定性混沌(而不是隨機性混沌),財政部長們遲早是無能為力的。部長會說,調節某個參量,好比說利率,對經濟會起長遠作用。可是別忘記「蝴蝶效應」:按照動力系統理論,長遠而論,一個領養老金的人把儲蓄從銀行取出來,也同樣可能導致經濟崩潰。
梅的論文使人們對生物群體的興亡有了一個新看法。可是,我們如果把目光從抽像數學模型的冷靜預言,轉移到諸如上面討論的山貓-野兔那種實際情況,我們就會發現,很難證明決定性混沌的確是觀測到的毫無規則的群體興亡的解釋。的確,儘管15年來的努力,還沒有任何人從生物群體研究中拿出一個得到公認的混沌實例來,其主要原因是數據不夠充分,無法給出一個明確的答案。
為了克服這個困難,有人設想了一些吸引人的實驗,來測驗非線性動力學的預言本領。其中一個是華盛頓大學應用數學家考特(Mark Kot)設計的,這個實驗和「無機化學」中的貝魯索夫-扎孛廷斯基反應十分相似。考特設計了一個捕掠一犧牲系統,其中捕掠者是一種名為原始動物的單細胞生物,犧牲品是某種細菌。億萬個這種有機體盛在一個開口的反應器中,細菌吃的食物不停流入,廢物不停提走。在考特的數學模型中,食物輸入如果穩定,這些微小有機體的群體動態就很簡單。但食物的供應如果模仿季節而做週期性循環,預言混沌就會出現,就像第六章的BZ反應一樣。目前進行的實驗,目的就是尋找這種混沌。
另一些近來的理論研究是倫敦皇家學院的安德森(Roy Anderson)和梅合作的。他們相信在有艾滋病的免疫系統中,他們找到了混沌的證據。這種系統裡可以說存在有一個捕掠-犧牲關係,艾滋病的來源——人體免疫缺乏病菌HIV「捕掠」白細胞和淋巴球T4,同時也刺激它們產生專門反抗HIV的免疫反應——B細胞與抗體。更廣泛地,流行病學家現在用混沌來分析麻疹、腮腺炎之類的蔓延。阿利桑那大學的沙費爾(William Schaffer)是這方面混沌最熱烈鼓吹者之一。沙費爾與其合作者分析了丹麥哥本哈根現有的流行病數據,自稱對麻疹、腮腺炎、風疹都取得了混沌的證據,而雞痘則有規則的週年循環。這些流行性疾病究竟是內在的混沌,還是外界隨機影響的結果,現在仍不清楚。
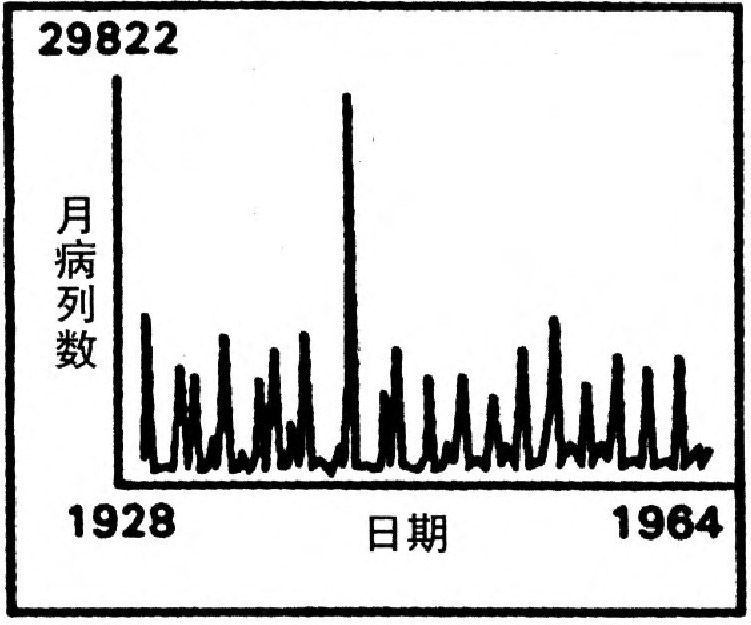
圖28 有序和無序的混合體:1928~1964年紐約城中的麻疹病例。
進化與深時
現在讓我們把話題轉回到進化上。1859年11月24日達爾文《經由自然選擇物種的起源》一書的出版,標誌了一個智慧革命,其影響超出生物學,徹底改變了有頭腦的人對世界的看法。達爾文的進化箭頭,從簡單的單細胞有機體指向複雜如人的生物,乍看上去是和波爾茲曼想解釋的熱力學箭頭相反。但有一點比什麼都重要,那就是:進化需要時間。地球存在的時間足夠長了嗎?
按照猶太-基督宗教的說法,地球創生後不到幾天,人就出現了。17世紀,北愛爾蘭阿爾瑪市的大主教鄂修(James Ussher)根據聖經,斷言世界創生於紀元前4004年。可是當地質學家們認識到化石是一度生活於地球上的生物的石化遺體,而地質層是按時間順序放置的,他們隱約體會到「深時」。
地質深時的概念從17世紀起,地位漸漸提高,到了19世紀初期,地球年齡被估計在100萬年到幾十億年之間。1863年,開爾文勳爵根據地球應該遵守的散熱率,做了一個著名的估計。他的假設之一是:地球一度曾處於熔態,然後漸漸冷卻;由此他得到結論,說地球支承生命不到十億年。隨後又有更精確的計算,把這可用的時間減到1000萬年。
19世紀末放射性的發現,徹底解決了地質時間尺度問題。地球中地核地幔之間的溫度梯度可能來自地殼中的放射性衰變,這樣,開爾文的時間尺度就完全不對了。隨後人們想到用放射性衰變的遺跡來測量地球本身的年齡。一個原子核自身的衰變率可以用來度量時間的流逝;這是一種很好的線型、非循環式時鐘。衰變率一旦已知,放射性遺跡和其產品之比,可以用來估計地球年齡。這樣,英國科學家斯特魯特(John Wil-liam Strutt,即日後的瑞利男爵)在1905年給出20億年的估計(用鐳的衰變),而美國的波爾特伍德(Bertram Boltwood)估計是22億年(用鈾的衰變)。
深時的發現產生了極大影響,但這個概念很難掌握。用哈佛大學的古爾德的話來說:「對深時做個抽像的、理性的瞭解並不太難——我知道1億是1後面放多少個零。但內心是否真正體會,那完全是另一回事。深時對我們說來太生疏了,我們只好打打比喻。我們高談『地質裡』,人類歷史只佔其中最後的幾寸地;或者闊論『宇宙年』,人類只是在爆竹迎新年前幾分鐘才出場。」古爾德認為麥克菲(John McPhee)在他《盆地與山脈》一書中用的比喻最生動。麥克菲要我們把地球歷史比做英國老長度單位——碼,這是英國某個國王從鼻尖到伸臂後指尖的距離。國王手指甲銼一下就把整個人類歷史銼掉了。深時——進化的奠基石,它的奧妙就在於此。
達爾文的理論
1809年,達爾文出生於英國西部希茹斯布利城。他在愛丁堡大學學醫,當時他覺得地質課「枯燥到令人難以置信……唯一對我起的作用就是使我下決心,一生再也不讀任何講地質的書」。他隨後考進劍橋的基督學院,準備將來做牧師,那時他「被裹入一群蕩徒之中,其中包括一些下流青年」。的確,他父親有一次對他說:「你什麼也不管,整天就是打獵,賽狗,捉老鼠,你將給自己丟臉,給我們家門丟臉。」
1831年12月27日,達爾文乘官船「獵犬號」揚帆啟程,於是開始了一個長達五年的環球航行,這航行將種下對有機世界一個新看法的種子。雖然達爾文此行是經由該船船長菲茲勞耶(Robert Fitzroy)的邀請,他們兩人之間關係並不好,再加上達爾文的暈船、生病,使達爾文麻煩重重。菲茲勞耶自信他會看相,能從一個人的面貌看出此人的性格:「像我這樣的鼻子,他非常懷疑我會有參加此行的足夠精力和毅力。」達爾文說。
1836年10月2日,達爾文回到英國海港法爾牟司。到此為止,他從一大批不同的地面、環境、動物、植物中,對他多年來猜疑的一個想法——動物植物經過長遠時期能演化為別種動物植物,已積累了確鑿的證據。但他直到20年後才發表這想法,而且還是因為一個英國年輕的自然學者華萊士(Alfred Wallace)的一篇論文。華萊士在一個熱帶島上害瘧疾發高燒時,靈感一現,獨立地想到自然選擇的概念。一如達爾文,華萊士一直也在思考著馬爾薩斯的人口論。華萊士寫道:「我當時在呆想這理論對任何物種的作用,適者生存這意念忽然閃入我腦中。」達爾文大吃一驚,找了兩個有影響力的朋友,組織講座,同時宣讀華萊士的論文和他本人1844年發表的一篇文章的摘錄。達爾文的文章先讀;雖然有人認為華萊士受欺,但華萊士本人卻承認這位長輩比他早。
達爾文由於他的「猴子論」,成為漫畫家挖苦的對象,且遭受科學家的嚴厲批評、宗教組織的強蠻攻擊。他的論點是:自從地球上有生命以來,物種從較簡單的形式演化成較複雜的形式,而自然選擇——最適應環境的物種遺存下來,不適應的被淘汰,為此演化提供解釋。這理論那時其實屬於假設,而不能看做一個對世界的一成不變的描述:當時還沒有任何確定性的證明,說這理論是對的。這理論的強處是它自稱比其他任何理論更符合觀測到的事實。以為達爾文式的演化一成不變是錯誤的;今日所有受尊重的進化派學者接受的都不是當初一字未動的原本。我們的知識一面在增加,這理論也就相應在演變,但就如梅所說:「所有這些研究都是在達爾文所樹立的牢固框架中進行的。」
達爾文給地球上生命的多種多樣形式下的配方,在現代原子、分子語言中,有了新的、更引人入勝的說法。在很大程度上,生命——至少在觀念上,即使不談它的細枝末節,可以從為其基礎的DNA分子的性質中,得到理解。上章後部討論生命起源時,已經遇到過這些分子。道金斯(Richard Dawkins)在他《自私的基因》一書中動人地描寫了DNA自我複製的能力。我們都是「殘存機器」,唯一的目的就是保護基因,就是要讓同一條DNA鏈,也就是讓這些巨分子所載的基因——決定我們面貌性格的藍本,更多的自我複製。分子生物學的大步進展是這生命分子圖案正確性的有力見證者。對生物內中的情況——小到每個分子,已取得了驚人的進步,得到的知識已用來設計了數代新藥品,改變了生物的基因從而設計了新的生物,為診斷疾病和遺傳性失調,設計了高度靈敏的檢測手段。
可是,就如我們一再陳述,單純的歸納主義也有缺點;如說今日的分子生物學有一點可批評,那就是:它複雜的枝節會把它的全面圖像遮蔽了。對個別分子太關心,會使我們忽略了更重要的、分子之間的相互作用和時間上的合作行為。這種合作性的非線性效應,在遠離平衡狀態之下戲劇性地出現,代表著生命構成的一個不可缺少的因素。
有人試圖用熱力學對演化做一宏觀層次上的解釋。布如克斯(D.Brooks)與懷利(E.Wiley)在他們頗有爭議的《演化與熵》一書中,說了一句中肯的話:「我們相信,大家都同意,有機體是處於遠離平衡狀態的耗散式結構。」對他們的方法說來不幸的是,他們混淆了不同的概念——第五章我們看到,自紐曼以來,信息論的概念和熱力學的熵一再被人混為一談。從熱力學的確可以搬過來的是不可逆性的重要性,時間之箭的重要性。
進化過程是不可逆的,這看法最早提出者之一是法國出生的道羅(Louis Dollo 1857~1931),1893年他發表鼎鼎大名的「道羅定律」。可是按照進化論大師之一、梅納德·史密斯(John Maynard Smith)的說法,自然選擇並不意味著時間上的有向性。道金斯的著作裡說道羅定律「常常跟進步是不可避免的這主觀主義的無稽之談混在一起,跟進化是違反熱力學第二定律這無知的廢話扯在一塊」。並沒有理由說進化過程中的一般傾向是不能倒轉過來的,他說:「如果在某段時期鹿角傾向於變大。很可能過一段時期以後,它又傾向於變小。道羅定律其實只是講,同一條演化途徑完全重走一次的統計概率是極小的……。道羅定律並沒有什麼神秘,它也不是我們要到自然界去『檢驗』的某種東西。它只是從概率的基本定理中得到的一個結果而已。」道羅定律中成問題的是它把不可逆性只放在「進行演化的系統」,而第二定律是講整體的,既涉及此演化系統,也涉及此系統的周圍環境(請回想第五章的討論)。
在英國沃裡克大學數學研究所曾任多年的所長,現為牛津哈特福特學院院長的塞曼(Christopher Zeeman),認為這兩個概念永遠不會扯在一起:「從第二定律談進化是不對的。模擬是在某個一定的,像物理這樣的層次做的。下一個層次可能是化學,然後依次是巨分子結構、細胞、生物、生態學,最後是進化。每個層次開頭都得用個不同的模擬。當你一口氣又講生命,又講進化,又講熱力學第二定律時,要知道它們之間根本沒有什麼關聯。」按照塞曼的看法,生物學不像物理學,它是沒有什麼普遍理論的:「物理學中的普遍理論,對生物學說來無用,要不然就是引力會影響胚胎成長這類的話。你要引入一大部機械,但它給不出什麼對生物學家有興趣的預言。當生物學家們面對一大套數學時,他們就要問:『這真正有用嗎?』總的說來,他們不會浪費時間學這套數學。」奧斯特爾說得更尖銳:「以為熱力學能告訴你生命如何演化,就等於以為把電視機放在熱量計裡就可以說明它如何工作一樣。」
另一方面,許多作者,包括梅納德·史密斯本人在內,都斷言過越來越高的複雜性是生物演化的一個標誌,這當然是指時間有向的一個過程。諾貝爾獎獲得者盧利亞(Salvador Lu-ria)恰當地描述了進化的性質:「進化,一如歷史,並不像擲錢幣或者一場紙牌戲。它具有另一種不可缺少的特徵——不可逆性。所有的未來都是現在的後裔,所有的現在都來自實際的過去,而不是來自可能的過去。人是現實的子孫,不是假設情況的子孫,而演化的現實——實際存在的生物,它們的範圍只不過是過去所有可能的機會中很小的一個樣本。」
美國一位古生物學家顧爾德(Stephen Gould)相信他在某些資料中找到了時間箭頭的證據。他研究的是有同一個現在已經不復存在的祖先的一些生物,它們在時間上的分佈。按照他的同事、弗吉尼亞科技學院的吉林斯基(Norman Gilinsky)所說:「歷史看上去是不對稱的。」顧爾德在演化樹的枝幹分佈中找時間的箭頭,其中樹枝的粗細代表物種數目的多寡。他發現在物種數目增加的同時,有機藍本的種類便在減少,枝幹分佈不對稱,頭輕腳重。對此他做如下解釋:早先是試驗階段,品種數目大增,隨後是規範階段,數目減少,有機體衰亡。顧爾德認為這是化石資料中最顯著的傾向。他寫道:「這傾向比其他能從世系變化中而得到的結論,都更清楚地給時間確定了一個方向。這傾向很可能也反映著一個對自然系統如何演變的更一般、更基本的規則。」
最低最低限度,我們也可以說熱力學的時間箭頭和進化論的是相互不衝突的。既然化學、物理、數學越來越明顯地賦予生物學活力,耗散式非線性動力學的概念也就很自然地滲入生物進化這課題。這是否對生物學帶來好處,只有讓時間來驗證了。然而,對一些人來說,他們已經抵抗不了自組織和混沌的誘惑,他們在進化時間中已經看到了熱力學第二定律中的同一個箭頭。
生物化學家皮考克(Arthur Peacocke)寫道:「這樣,宇宙表面上的腐朽、趨向無序的傾向,為新形式的產生提供了必需的、不可缺少的母體(妙詞!)——經過老的死亡腐爛出現新的生命。」隨後又寫道:「普裡高津和艾根與其合作者的研究揭示出,在導致有機結構出現的過程中,機遇和法則(即必然)之間,隨機性與決定性之間的相互作用會是何等的微妙。他們的研究證明機遇和法則之間的相互作用其實具有時間上的創作性,因為是它們兩者之間的結合讓新的形式出現、演化——的確,自然選擇看上去是投機取巧的。這機遇和法則之間的相互作用,其性質現在看起來是使有機結構不得不產生,不得不演化。」
主張生物學中運用熱力學的人認為熱力學提供的是整個圖像的輪廓,而不是細枝末節,是可以借助動力系統理論把其他概念掛在上面的框架。情況和研究無機體中的自我組織時——BZ反應以及其他許多生物化學過程,完全類似:傳統的熱力學向不可逆非線性動力學讓步。例如,皮考克與其他人一起,斷言不可逆熱力學「可以用來淘汰某些生物學模型,因為它們跟宏觀物理定律相衝突,也可以容許一些模型,即使它對模型不能加以選擇」。
演化的步伐
演化時間之箭是枝斷箭:如果我們把所有現有的化石按照時間排列,它們並不像電影中連續的幀幅,每幀到下一幀的變化幾乎看不出來,而是似乎含有不連續的跳躍。上帝創生派和其他極端的原教旨派先用連續的演化箭頭醜化了進化論,然後說這證明進化論是錯誤的。
化石資料當然是不完整的。但資料中這些空缺也可能對所謂「間斷平衡」提供證據。「間斷平衡」這詞是艾爾德利基(Niles Eldridge)和顧爾德在1972年提出的。他們認為,根本不存在一個固定的演化率。物種傾向於在悠長的時間中保持穩定;它如要演化,演化就在很短暫的期間內進行。實際上,化石資料是由一層一層很厚的大陸土壤組成,每一層裡面物種均勻分佈,而層與層之間是物種突然變化的「面」。在這樣一個「間斷」點,一個物種一般演化成多個新種,如下面圖29所示。
達爾文理論和艾爾德利基-顧爾德理論之間有矛盾嗎?沒有。塞曼證明了達爾文式的演化能解釋間斷平衡這現象。塞曼對這些演化跳躍的分析基於湯姆(Ren Thom)的「災變理論」,該理論目的是:處理連續的「因」產生非連續的「果」的情況。達爾文式演化——隨機性微小變異和自然選擇,一翻譯成最簡單的數學形式,結果不是別的,就是圖29所示的間斷平衡。塞曼寫道:「我把達爾文的話翻譯成最簡單的數學函數,然後環境的變化就預言這種間斷平衡。」有趣的是:達爾文在其《物種的起源》一書中,唯一用以說明作者心意的插圖,就是圖29。

圖29 多叉圖、[錄自E.塞曼《達爾文式演化的動態學》一書。]
儘管他的模型取得了成功,塞曼對數學在演化理論中可能扮演的角色,頗講求實用。他相信,由於生物演化理論的很大一部分本質上始終將是描述性的,數學扮演定性的角色比扮演定量的角色更為可能。然而:「正是在與直覺相違背、用語言難以說服,例如連續的前因產生非連續的後果這類的場合,數學可以期望起作用。」
混沌:性別存在的理由
演化論另一難題是性別,這問題可以用決定性混沌在時間之箭中的出現來對付。我們周圍許多種生物是有性生殖。有些像細菌之類是無性生殖,那就是說,一個細菌不要另一個細菌幫忙,就能製造自己的複製品。還有別的,例如有翼小昆蟲薊馬,它們兩樣都來。可是,為什麼性別會演化出來?對進化最幼稚的看法是:基因是否被選,全看它的殘存母體是否在為有限資源的戰鬥中佔到上風。適應不夠的就死亡。可是,梅納德·史密斯在他關於進化與有性生殖的繼續保存的一本專著中,宣稱對性別的瞭解,目前仍缺少某個基本因素。使他感到困惑的是:短期看來,無性生殖更佔優勢。
有性生殖具有一些明顯的缺點:找個對象會很麻煩;不同個體的基因聚集到同一個後裔,每個個別個體的基因總量只傳了一半。這看上去似乎跟「自私基因」概念衝突。一個有性生殖的雌性動物平均每生一個雌性後裔時,無性生殖的動物已經生了兩個;因此在兩種都有的情況下,無性動物很快地會佔上風。
可是性別也有它的優點,即使不算伍迪·阿侖(Woody Allen)所說的「這是我不笑而覺得最好玩的東西」。優點肯定是有的,不然我們也就不在這兒討論這問題了。有性生殖比無性生殖更有效地把基因混合,使群體能有更多的遺傳變異。這伸縮性的增大使群體更容易適應環境的變化。並且在短期內,有性生殖使群體更容易對付新興的敵人,例如過濾性細菌,因為它使敵方更難預測。我們知道許多有性生殖的群體,它們比無性群體更能抵抗寄生物的入侵;例如,引入智利的一種銹菌大大減少了一種無性生殖的黑莓(學名Rubus constrictus)的成長,而對有性生殖的黑莓(Rubus ulmifolius)則影響幾無。
決定性混沌能幫我們瞭解,性別為什麼在自然選擇的過程中出現,使得我們更能對付寄生物。這至少是牛津大學動物系的哈密爾頓(Bill Hamilton)和他同事的觀點。他們研究了性別在一個演化遊戲中起的作用,其中相互競爭的宿主盡力設計對付寄生物的策略。在這計算機上進行的鬥爭中,哈密爾頓模擬了宿主和寄生物的遺傳構造。簡單說來,寄生物的目的是使自己與宿主的遺傳構造相配合,從而撲向宿主,把它吃掉。遊戲裡,配合得越好,寄生物的「適應分數」就越高,它就越成功。配合完全,宿主遭殃。反過來,宿主盡量使自己的遺傳構造和寄生物的「不配合」,使自己更容易避開寄生物。哈密爾頓的數學模型用13個耦合非線性方程來顯示群體在時間上的演化。他認為,如對引入模型中的參數做現實的選擇,結果將是嚴格意義下的決定性混沌。
按照哈密爾頓的看法,這裡如果出現一個奇異吸引子,將對性別有利,雖然此點尚未確證。原因是:奇異吸引子帶有混沌式的、不可預測的演化,有這一手,一個有性生物總比其寄生物佔先一步。一個行為受其基因影響很大的生物(當然環境也起作用),混沌比光靠自然變異對演化提供更大的不可預測的餘地。
哈密爾頓認為,混沌不是性別存在的必然原因;只是性別如果存在,經由自然選擇的長期演化就更順利。和單性生殖相比,兩性生殖使生物更快地駛入混沌安全港。哈密爾頓並不是混沌的「傳教士」,他談起自己運用不可逆非線性動力學,態度頗為輕蔑;他說:「我的做法其實很笨。我只是把它放進計算機,然後看出些什麼。」應該指出,像性別這本身惹人爭論的課題,關於它為什麼會演化出現,目前還有不少其他的建議。
生物界的節拍
在許多方面,地球上的生命像一支交響樂團,按照上天的節拍奏樂。太陽、月亮的運行,反映在昆蟲總數的起伏之中,反映在全球有生命物體的活動之中。一切都是時間上的圖案。
在用以描述這些圖案的產生的理論框架裡,時間總是向同一個方向走。在這枝時間之箭上刻著無止境的變化。有些圖案人眼可以看到,例如金錢豹的點紋,戚納巴魯金蘭的條紋。有些看不見,例如心臟的跳動,神經細胞的激發,細胞分裂的不斷進行。這些時間上的圖案不僅是生命的一部分,而且是生命的基礎。即使表面看上去是偶然性的過程,諸如疾病的傳染,魚群總數的漲落等,裡面可能都隱藏著秩序。不可逆非線性動力學裡面包含既有自組織的配方,也有動力學混沌的配方。
宇宙在時間上普遍地、單向地前去,朝著一個最大熵的可能狀態,在這過程中,滔滔湧出細巧有序而瞬息即逝的生命圖案。否定這時間之箭,對諸如相對論、量子力學、經典力學等領域說來,或許是方便的——話說回來,這些理論適當運用在別的場合是相當成功的。但也許這些巍然的理論建築未能說明事物的全部真相。
法國理論物理學家德·艾斯帕納(Bernard d'Espagnat)曾說:「從諸如災變、耗散式結構等概念,到對有機物體的演化一個真正普遍的瞭解,這中間仍是相當大的一步。……但是這方面確有進步,這是不可否認的。希望將有更多的進展,早晚會漸漸看到出現一套理論,它非但不把『活力』歸並於某些平常的機制,並且要把『活力』深奧的美,更清楚地揭示給我們看,就像經典天文學把宇宙的美揭示給我們先人一樣。」